Evidence for Negative Frequency Dependent Sexual Selection on Eye Color in Europeans
Abstract
Human populations exhibit a variety of eye color phenotypes, with Europeans exhibiting more diversity than others [1-3]. A variety of selective mechanisms to explain this pattern have been proposed; these include a general preference for blue eyes [4,5], parental imprinting effects [6-8] inbreeding avoidance behaviors [9,10], paternity assurance preferences [11,12] and frequency dependent selection favoring rare phenotypes (termed negative FdSs) [1]. Here we show, using an ethnically diverse study population asked to rate the attractiveness of faces of Caucasian members of the opposite sex in which the eye color frequencies were manipulated, strong support for negative FdSs exhibited by white females of European ancestry. Our data shows no support for FdSs in females of other ethnicities or males and a lack of support for any of the other models listed above. These results are consistent with a model of human evolution in which a history of negative FdSs by European females, but not males, is responsible for the high degree of eye color diversity seen in modern Europeans.
Keywords
Sexual selection, Frequency dependent, Eye color
Introduction
Human populations exhibit a variety of eye color phenotypes, with Europeans exhibiting more diversity than others, especially for blue and green colors [1-3]. The ancestral eye color state in Europeans was brown, with the blue and green colors recently derived within the past 6-10,000 years [1-13]. Although not as simple as a multi-allelic one locus trait, the number of genetic factors controlling the blue-brown eye color variants appear to be few in number [2,3,14,15] with green eyes arising as an intermediate phenotypic state. Studies report that blue eye color is associated with either no difference in visual acuity [16] or reduced quality of vision [17] which suggests that the increase in frequency of blue eyes likely arose from sexual selection, which has long been accepted as a powerful evolutionary force [18-20].
Previous studies of sexual selection in humans have identified a number of factors that influence ratings of attractiveness and mate choice [21-24]. Surprisingly, very few studies have examined eye color preference in humans. Although subjects report a preference for blue eyes over brown when asked [4,5], when preferences were actually tested by having subjects rate the attractiveness of eyes or faces with digitally manipulated eye colors there was no overall difference in attractiveness ratings given to individuals, faces or eyes exhibiting different eye colors [5,11,25,26] or brown eyes were actually preferred over blue [12]. However, due to the relatively low number of studies, the existence of an overall preference for certain eye colors remains unknown. Sexual selection based on eye color could also arise from imprinting effects favoring individuals with the same color eyes as the opposite sex parent [6-8]. Conversely, selection for behavioral factors that minimize inbreeding by creating preferences against the eye color of siblings, which are necessarily correlated with that of their parents, via the "Westermarck effect" has been proposed [9,10]. Furthermore, because kin recognition systems may be self-referential rather than family-referential [27], selection for or against eye colors similar to oneself may arise as an extension of the aforementioned imprinting processes. Additionally, blue-eyed males may prefer blue-eyed females due to paternity assurance benefits [11,12] or because a general preference for self-similarity may exist [12,28,29]. Selection processes favoring certain colors and positive assortative mating and will lead to reduced genetic variation and fixation of favored phenotypes instead of polymorphism however and do not provide a mechanism for the increase of the genetic factors responsible for blue and green eye colors in Europe and subsequent maintenance.
If preferences and fitnesses are determined by frequency, with rarer phenotypes favored, then this negative frequency dependent sexual selection (negative FdSs) can act to both increase the frequency of initial mutations and maintain phenotypic variation in populations for prolonged periods of time [30-32]. Negative FdSs on eye color has therefore been proposed as the mechanism by which blue and green eye colors first became frequent and have since been maintained in Europe [1]. More precisely, the proposed hypothesis was one in which, during a prolonged period of female-skewed sex ratios during the period approximately 10,000 years ago, sexual selection by males for females with rare eye color variants led to the present color polymorphism [1]. This hypothesis has not been directly tested.
Methods Summary
The study was conducted during the period from summer 2011 to spring 2012 using students at California State University Long Beach. Internal CSULB ethics clearance was obtained for this study. Subjects were presented with altered photographs of the faces of Caucasian members of the opposite sex and were asked to rank order the attractiveness of images displayed in sets of six (an example of a set is shown in Figure 1). The eye colors of the displayed images had been altered to randomly display as either blue, brown or green and the frequencies of the two eye colors displayed within each of the five sets displayed to each subject were manipulated (1:5, 2:4, 3:3, 4:2, and 5:1). Different subjects therefore ranked the same face over a range of circumstances in which that face possessed each eye color as a common, intermediate or rare variant. Subjects then completed confidential questionnaires in which they provided their sex, ethnicity, age, eye color and parents' eye colors. Data for 366 subjects passed quality control procedures with sufficient representatives from White, Asian and Hispanic ethnicities for separate analyses of members of these ethnic subgroups (no other single ethnic category had sufficient representation to conduct separate tests). Statistical analyses were conducted using the One-Way ANOVA and General Linear Model modules in Minitab 16 and via simulation studies. Forms and surveys provided to subjects, summaries of all statistical results and the computer code used for the simulation analyses are available in the Supplementary Materials.
Methods
Study participants
This study was conducted using student participants, primarily undergraduates, at the California State University Long Beach (CSULB) during the period from summer 2011 to spring 2012. Internal CSULB ethics clearance was obtained for this study. On the day of participation, subjects were given informed consent by one of the study authors and then completed the study. The study had two portions: an assessment of faces presented to the subject and a biographical questionnaire in which subjects recorded their sex, ethnicity, age, eye color and parents' eye colors. Data from subjects who made errors in the assignment of rankings, recognized any faces or were older than 30 were removed from the analysis which resulted in data from 366 subjects remaining for analysis. A sufficient number of representatives from White, Asian and Hispanic ethnicities were present for analyses of data within each of these ethnic subgroups (no other single ethnic category had sufficient representation to conduct separate tests).
Study design
Each subject was seated at a computer and they were directed to a password-secured website where, after entering their sex, they were presented with 5 sets of 6 digitally altered images. The images were the faces of Caucasian individuals of the opposite sex and the subjects were asked to rank the attractiveness of the six faces within each set on a scale from most attractive to least attractive (1-6 with no duplicated values allowed). Subjects were given written directions and an assessment form (Supplementary Materials files S1 and S2) on which to record their rankings which included a code number to ensure their anonymity, but allowed these forms to be matched to confidential biographical questionnaires they completed later.
Each of the 5 sets of faces displayed the same individuals, but the images were digitally altered such that the eye colors were set to blue, brown or green using Photoshop™ (an example of a set is shown in Figure 1). All subjects of each sex were exposed to the same 30 faces and the general shapes and/or attractiveness of the faces were subjectively chosen to be similar within each set of six. The 5 sets were presented in random order and within each set all faces displayed one of two selected eye colors randomly chosen such that each of the 3 pairwise color combinations was used once and the remaining two sets used a randomly determined pair of colors. The 5 sets displayed each of the five eye color frequencies available with six faces (1:5, 2:4, 3:3, 4:2, and 5:1) randomly. After ranking the faces in order of attractiveness, subjects completed the biographical questionnaire (Supplementary Materials file S3).
Statistical analysis
A full set of the statistical results and the C++ program used to perform the simulations described below is available in the online Supplementary Materials file S4.
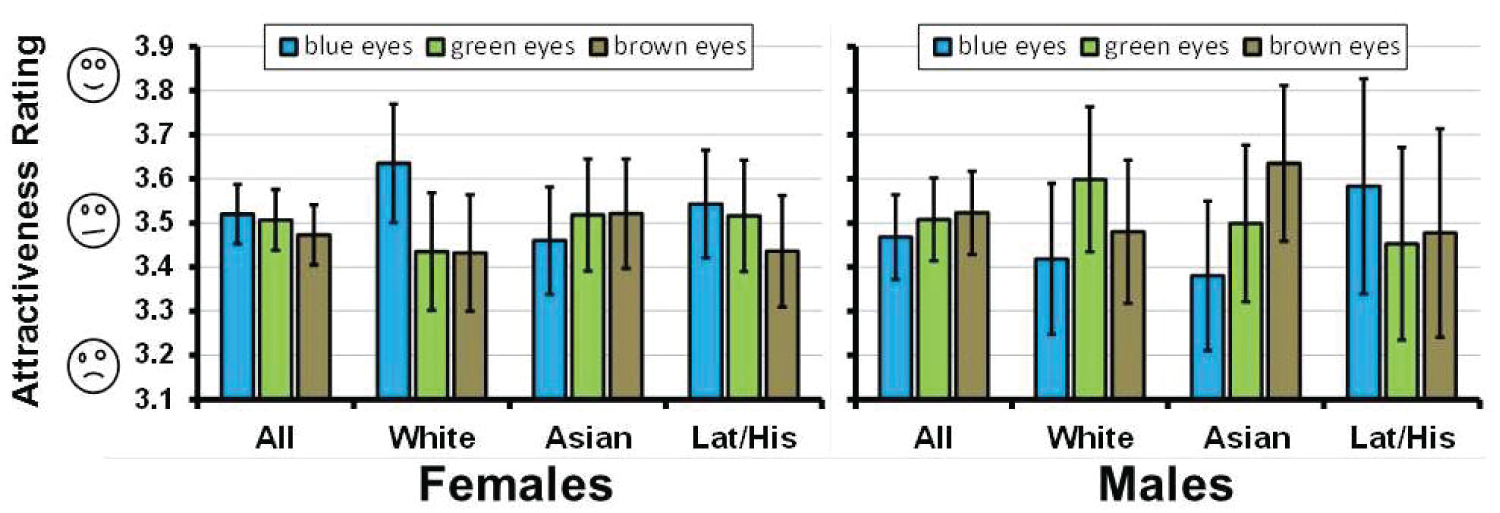
Differences in mean attractiveness rankings for faces exhibiting different eye colors (see Figure 2) were examined using the One-Way ANOVA module in MiniTab 16 with eye color as the categorical predictor variable.
Evidence of patterns consistent with imprinting or paternity assurance in White subjects was examined using the General Linear Model module in MiniTab16. Separate unbalanced Two-Way ANOVA analyses of attractiveness ranking using study subject eye color, same sex parent eye color and opposite sex parent eye color in combination with photograph eye color were performed. The subjects used for each analysis necessarily differed from analysis to analysis due to the differential omissions of subjects reporting eye colors not falling into the three categories analyzed (e.g., hazel) for themselves or either parent.
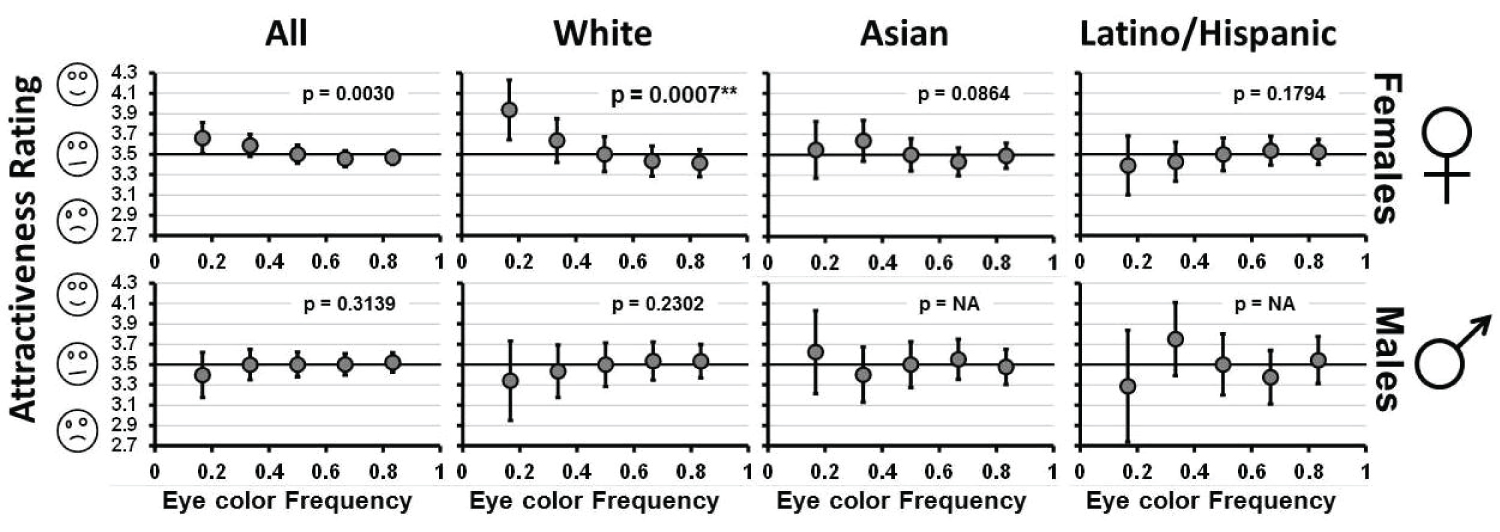
The study design inherently creates a non-linear relationship between frequency and mean ranking because each rank is used once; the possible range of mean values are therefore more constrained for more common eye colors than for rare ones and a linear regression analysis is inappropriate. We therefore performed a series of simulations in which simulated subjects provided rankings randomly and the proportion of trials from 10,000 performed in which the mean attractiveness rankings were more extreme than that observed for both the rare frequencies was recorded (i.e., this is the p value). The number of simulated subjects was set to match the number in each sample group and the two-tailed nature of the test allows detection of patterns consistent with both negative and positive FdSs. We consider data in which the two rare frequencies showed a mixed pattern (i.e., one above 3.5 with the other below) to be indicative of no overall FdSs and testing with this technique was deemed non-informative (i.e., p = NA). The analysis was performed on all individuals within each sex and separately for each of the 3 subsets of the study subjects defined by their self-reported ethnicities (White, Asian/Asian American, Latino/Hispanic) and for a set consisting of all non-White females.
We therefore tested for eye color preferences, the presence of imprinting and paternity assurance effects and patterns of FdSs by asking study subjects to rank the attractiveness of faces in which the eye colors had been digitally manipulated to be blue, brown or green (see Figure 1). Sets of six faces were presented to the subjects and all the faces in each set had one of two eye colors with the frequencies of the eye colors varied such that the same face would exhibit the rare, moderate, or common eye color randomly when presented to different subjects. The diverse ethnic and gender makeup of the subjects allowed separate analyses of subgroups of individuals self-reporting as White, Asian/Asian-American and Latino/Hispanic for both males and females. The eye colors of the subjects and their parents were recorded and for the White subset of our data we tested the imprinting and paternity assurance hypotheses described above.
Results
Several of the factors previously proposed to cause preferences for eye color were not apparent in our data. Although some faces were consistently rated more attractive than others, none of the eye colors showed a significant difference from the others in mean attractiveness in the overall sample or within each of the ethnic subsets (Figure 2). This result is consistent with previous reports showing no preference for blue eyes even by those that claim a preference for blue eye color [5], but is in contrast to previous studies showing a stated preference for blue [4,5] or measured preference for brown eyes [12].
The subset of White study subjects was used to test the parental imprinting and Westermarck hypotheses in both females and males. No meaningful significant interactions were seen between the attractiveness of faces with certain eye colors and the eye color of the subject or either parent in any of the tests (see online supplementary materials). In a more focused analysis, the subset of blue and brown-eyed male study subjects was used to test the paternity assurance hypothesis for mate choice by examining their rankings for blue and brown eyed faces. No significant interaction between eye color and preference for eye color was seen in the subset of blue and brown eyed male White study subjects and the non-significant preference for faces with blue-eyes over faces with brown eyes was actually less pronounced in blue-eyed males than in brown-eyed males (see online Supplementary Materials).
Preference for eye color did appear to be significantly related to frequency however. The relationships between eye color frequency and attractiveness ranking are shown in Figure 3. There is evidence for significant negative FdSs in which females favor male faces with rare eye colors in the overall data set (p = 0.003 see online supplementary materials), but this pattern is driven almost entirely by the highly significant negative FdSs relationship observed in the White subset of individuals (p = 0.0007, see online supplementary materials). The Asian and Hispanic subsets of subjects and a data set consisting of all non-White females exhibited no significant relationship between eye color frequency and attractiveness ranking. There was no significant evidence of FdSs in males in the overall data set or within any of the ethnic subsets.
Discussion
Our data therefore provides no evidence of overall preference for certain eye colors, imprinting effects, a Westermarck effect or preferences related to paternity assurance, but does provide strong evidence for negative FdSs in the choices made by females of European ancestry. In contrast, males of European ancestry did not show evidence of FdSs and the non-significant trend evident is the opposite of that predicted by a model of negative FdSs in choices made by males [1]. Evidence for choice in females, but not in males is consistent with the observation that females generally exhibit higher degrees of mate selectivity than males across a wide range of species [19,20,33].
Our results support a model of human evolution in which a history of negative FdSs by European females, but not males, is responsible for the high degree of eye color diversity seen in modern Europeans. The lack of evidence for negative FdSs behaviors in the other ethnic groups we studied is consistent with the observed pattern of global eye color diversity and suggests that the lower eye-color diversity seen elsewhere is due to a historical lack of negative FdSs in those regions. We also note that the ethnic subset comparisons that support our conclusion were made possible by using a diverse study population that showed evidence of distinct preferences; a lack of such diversity has been noted as a weakness in many psychology studies [34,35], and we therefore encourage similar studies in the future to actively incorporate members from different ethnicities.
Supplementary Information
Supplementary Information is linked to the online version of the paper
Acknowledgements
We thank J. Uhl for design of the website used; D. Jones for photographic assistance; A. Salomon, N. Fitch and S. Sweeting for data recording and checking; and M. Young-Owl and C. Warren for critical discussions. We dedicate this paper to the memory of J. A. Papagno.
Author Contributions
A.J.R.C. designed the study, conducted the study and analyzed data. A.U. took and edited the photographs, conducted the study and contributed to writing the paper. A.J.R.C. and A.U. wrote the paper jointly. All authors discussed the results and commented on the manuscript.
Author Declaration
The authors declare no competing financial interests.
Summary for Journal Readers
Humans exhibit a much higher variety of eye colors in European populations than elsewhere. Sexual selection for traits based on their frequency (e.g., rare phenotypes are favored) may explain this variation. We found that Caucasian female subjects rated male faces as more attractive when they exhibited relatively rare eye colors than when their eye color was common, but we did not see this pattern in females of other ethnicities or males. This pattern is consistent with female choice driven sexual selection as seen in many wild animals and may explain the higher eye color diversity seen in Europeans.
Summary for the General Public
Why do Europeans tend to have more eye colors than non-Europeans? One hypothesis is that historical selection for individuals with rare eye colors has maintained many diverse eye color genes by favoring each when they get rare. This study shows that young Caucasian females seem to prefer certain male faces more when they have relatively rare eye colors whereas females of other ethnicities and males don't show the same bias. A long history of preferences like these, rather than any preference for a single color, may explain why Europe currently has more eye color diversity than other regions.
References
- Frost P (2006) European hair and eye color: A case of frequency-dependent sexual selection? Evolution and Human Behavior 27: 85-103.
- Sulem P, Gudbjartsson DF, Stacey SN, et al. (2007) Genetic determinants of hair, eye and skin pigmentation in Europeans. Nat Genet 39: 1443-1452.
- Eiberg H, Troelsen J, Nielsen M, et al. (2008) Blue eye color in humans may be caused by a perfectly associated founder mutation in a regulatory element located within the HERC2 gene inhibiting OCA2 expression. Hum Genet 123: 177-187.
- Feinman S, Gill GW (1978) Sex differences in physical attractiveness preferences. The Journal of Social Psychology 105: 43-52.
- Grundl M, Knoll S, Eisenmann-Klein M, et al. (2012) The blue-eyes stereotype: Do eye color, pupil diameter, and scleral color affect attractiveness? Aesth Plast Surg 36: 234-240.
- Bereczkei T, Gyuris P, Koves P, et al. (2002) Homogamy, genetic similarity, and imprinting; parental influence on mate choice preferences. Personality and Individual Differences 33: 677-690.
- Bereczkei T, Gyuris P, Weisfeld GE (2004) Sexual imprinting in human mate choice. Proc Biol Sci 271: 1129-1134.
- Little AC, Penton-Voak IS, Burt DM, et al. (2003) Investigating an imprinting-like phenomenon in humans partners and opposite-sex parents have similar hair and eye color. Evolution and Human Behavior 24: 43-51.
- Westermarck E (1891) The History of Human Marriage. Macmillan, London, UK.
- Rantala MJ, Urszula M (2011) The role of sexual imprinting and the Westermarck effect in mate choice in humans. Behav Ecol Sociobiol 65: 859-873.
- Laeng B, Mathisen R, Johnsen J-A (2007) Why do blue-eyed men prefer women with the same eye color? Behavioral Ecology and Sociobiology 61: 371-384.
- Bovet J, Barthes J, Durand V, et al. (2012) Men's preference for women's facial features: Testing homogamy and the paternity uncertainty hypothesis. PLoS One 7: e49791.
- Cavalli-Sforza LL, Menozzi P, Paizza A (1994) The history and geography of human genes. Princeton University Press, Princeton.
- Mengel-From J, Børsting C, Sanchez JJ, et al. (2010) Human eye colour and HERC2, OCA2 and MATP. Forensic Sci Int Genet 4: 323-328.
- White D, Rabago-Smith M (2011) Genotype–phenotype associations and human eye color. J Hum Genet 56: 5-7.
- Short GB (1975) Iris pigmentation and phototopic visual acuity: A preliminary study. Am J Phys Anthropol 43: 425-434.
- Nischler C, Michael R, Wintersteller C, et al. (2013) Iris color and visual functions. Graefes Arch Clin Exp Ophthalmol 251: 195-202.
- Darwin C (1871) The Descent of Man, and Selection in Relation to Sex. John Murray, London, UK.
- Andersson M (1994) Sexual Selection. Princeton University Press, Princeton, NJ.
- Andersson M, Simmons LW (2006) Sexual selection and mate choice. Trends in Ecology & Evolution 21: 296-302.
- Kościński K (2007) Facial attractiveness: General patterns of facial preferences. Anthropological Review 70: 45-79.
- Rhodes G (2006) The evolutionary psychology of facial beauty. Annu Rev Psychol 57: 199-226.
- Perrett D (2010) In Your Face: The New Science of Human Interaction. Palgrave Macmillan, London.
- Swami V, Salem N (2011) Evolutionary psychology: A Critical Introduction. Swami V, 131-182, Wiley-Blackwell, 2011.
- Hickson M III, Powell L, Sandoz ML (1987) The effects of eye color on homophily, attraction, and credibility. Communication Research Reports 4: 20-23.
- Kleisner K, Kočnar T, Rubešová A, et al. (2010) Eye color predicts but does not directly influence perceived dominance in men. Personality and Individual Differences 49: 59-64.
- Bressan P, Zucchi G (2009) Human kin recognition is self- rather than family-referential. Biol Lett 5: 336-338.
- Little AC, Burt DM, Perrett DI (2006) Assortative mating for perceived facial personality traits. Personality and Individual Differences 40: 973-984.
- Kocsor F, Rita Rezneki R, Szabolcs Juhász S, et al. (2011) Preference for facial self-resemblance and attractiveness in human mate choice. Arch Sex Behav 40: 1263-1270.
- Thelen TH (1983) Minority type human mate preference. Soc Biol 30: 162-180.
- O'Donald P, Majerus ME (1988) Frequency-dependent sexual selection. Philos Trans R Soc Lond B Biol Sci 319: 571-586.
- Gray SM, McKinnon JS (2007) Linking color polymorphism maintenance and speciation. Trends Ecol Evol 22: 71-79.
- Clutton-Brock T (2007) Sexual selection in males and females. Science 318: 1882-1884.
- Henrich J, Heine SJ, Norenzayan A (2010) Most people are not WEIRD. Nature 466: 29.
- Henrich J, Heine SJ, Norenzayan A (2010) The weirdest people in the world? Behavioral and Brain Sciences 33: 61-135.
Corresponding Author
Alexandra Uhl, California State University Long Beach, USA
Copyright
© 2023 Uhl A, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.