The Discovery of the "Calcium Paradox" Due to Ca2+/Camp Interaction: Novel Adventures for the Pharmacotherapy of Neurological/Psychiatric Disorders
Our discovery of the involvement of the interaction between intracellular signalling pathways mediated by Ca2+ and cAMP (Ca2+/cAMP interaction) in the neurotransmission and neuroprotection has produced new avenues in the understanding of the cellular and molecular mechanisms involved in the pathogenesis of neurological and psychiatric disorders, such as Alzheimer's and Parkinson's diseases. Interestingly, this discovery initiated decades ago when numerous clinical studies have reported that use of L-type Ca2+ channel blockers (CCBs) by hypertensive patients decreased arterial pressure, but produced typical symptoms of sympathetic hyperactivity, such as tachycardia and increment of catecholamine plasma levels. Despite these adverse effects of CCBs have been initially attributed to adjust reflex of arterial pressure, during almost four decades this enigmatic phenomenon named "calcium paradox" remained unclear. In 2013, we discovered that this phenomenon was resulting of increment of transmitter release from sympathetic neurons and adrenal chromaffin cells, stimulated by CCBs due to its interference on the Ca2+/cAMP interaction. In this way, our discovery of the role of Ca2+/cAMP interaction in the neurotransmitter release, and neuronal death triggered by cytosolic Ca2+ overload, opened novel adventures for the development of new pharmacological strategies more effective for the treatment of neurological and psychiatric disorders resulting of neurotransmitter release deficit, and neuronal death.
Keywords
Ca2+/cAMP interaction, Calcium paradox, Neurological/Psychiatric disorders
Introduction
Numerous results which have been originated decades ago, using chromaffin cells as secretory model, established the notion of stimulus-secretion coupling to explain neurotransmitter release. This notion was initially resulting from the experiments performed by Douglas and Rubin in the 1960s to study acetylcholine-stimulated secretory response in cat adrenal gland [1]. Using adrenal chromaffin cells, Baker and Knight revealed in 1970's that a rise in the cytosolic Ca2+ concentration ([Ca2+]c) is an elementary requirement to trigger transmitter release [2]. The demonstration of direct relationship between rapid neurotransmitter release and rise in [Ca2+]c derived from the experiments using photo released caged Ca2+ in adrenal chromafin cells performed Neher and Zucker in 1990's [3]. Thus, it has been classically accepted that Ca2+ has an important role in several cellular responses, notable by controlling neurotransmitter release and smooth muscle contraction. Putatively, the use of L-type Ca2+ channel blockers (CCBs) should reduce these cellular responses. In addition to Ca2+, many results have shown that cAMP increases neurotransmitter release at many synapses in autonomic nervous system of vertebrate, including sympathetic neurons [4]. Although the cellular and molecular mechanisms involved in these facilitatory effects of cAMP on the release of neurotransmitter and hormones are indistinct, the evidences suggest that this important intracellular messenger modulates intracellular signalling mediated by Ca2+ involved in the regulation of neurotransmitter, and hormones release.
Implications of the Ca2+/cAMP Signalling Interaction in Neuronal Function
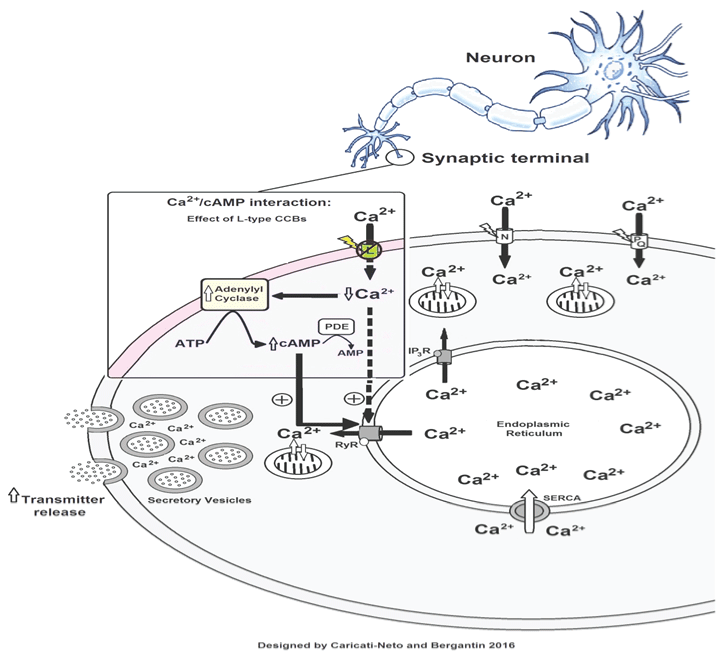
In fact, the hypothesis for a suitable interaction between the intracellular signalling pathways mediated by Ca2+ and cAMP, named Ca2+/cAMP interaction, has been widely studied in different cell types and tissues. In general, this interaction results in synergistic actions of these intracellular messengers on cell functions regulated by adenylyl cyclases (ACs), or phosphodiesterases (PDEs) [5-8]. The Ca2+/cAMP interaction has particularly been extensively studied at the endoplasmic reticulum (ER) Ca2+ channels, such as Ca2+ channels regulated by ryanodine receptors (RyR) [5-8]. Our studies established that Ca2+/cAMP interaction plays a role in neurotransmitter release regulation in neurons and neuroendocrine cells [5-8]. Then, dysfunctions of cellular homeostasis of Ca2+ and/or cAMP in these cells could result in the dysregulation of Ca2+/cAMP interaction, and could be a novel therapeutic goal for medicines.
Pharmacological Manipulation of the Ca2+/cAMP Interaction and its Consequences in Neuronal Function and Neuroprotection
Considering that Ca2+ has been classically implicated in several cellular responses, for example by controlling neurotransmitter release and smooth muscle contraction. Then, the use of CCBs should reduce these cellular responses. However, since four decades ago, several medical studies have been evidencing that acute and chronic use of CCBs in the antihypertensive therapy, such as nifedipine and verapamil, decreased peripheral vascular resistance and arterial pressure arterial, but produced typical symptoms of sympathetic hyperactivity such as tachycardia, and increment of catecholamine plasma levels [9]. Despite these adverse effects of CCBs have been initially attributed to adjust reflex of arterial pressure, during almost four decades the cellular and molecular mechanisms involved this enigmatic phenomenon named "calcium paradox" remained unclear. In addition, besides the role of Ca2+ in controlling neurotransmitter release and smooth muscle contraction, an imbalance of intracellular Ca2+ homeostasis has been implicated in the pathogenesis of aging-related neurodegenerative diseases. Several evidences suggest that aging impairs ability of the brain intracellular Ca2+ degradation, which is likely to induce cellular damage due to cytosolic Ca2+ overload, leading to neural death and resultant cognitive dysfunction, such as Alzheimer's disease [10]. From this concept in mind, a 10-year follow-up study (2000 to 2010), involving 82,107 hypertensive patients of more than 60 years of age, showed that use of L-type CCBs reduced blood pressure and risk of dementia in hypertensives, suggesting that these drugs could be clinically used to treat Alzheimer's disease [11]. Supportive findings for the neuroprotective effects of CCBs have been demonstrated in 1,241 elderly hypertensive patients with memory impairment [12]. The use of CCBs decreased the risk of cognitive impairment, and Alzheimer's disease, independently of blood pressure levels when compared to patients not receiving CCBs [12]. The long-term effects of antihypertensive therapy, initiated with a long-acting dihydropinidine (nitrendipine), have been demonstrated in the double-blind, placebo-controlled Syst-Eur trail in which the incidence of dementia was reduced by 55% [13].
Returning to the enigma of the "calcium paradox", we discovered about 4 years ago that the "calcium paradox" phenomenon was resulting of increment of transmitter release from sympathetic neurons, and adrenal chromaffin cells, stimulated by CCBs due to its interference on the Ca2+/cAMP interaction. Using isolated tissues richly innervated by sympathetic nerves (rat vas deferens) to exclude the influence of adjusting reflex, we showed that neurogenic responses of the vas deferens were completely inhibited by L-type CCBs in high concentrations (> 1 μmol/L), but unpredictably, and paradoxically, potentiated in concentrations below 1 μmol/L, characterized by sympathetic hyperactivity induced by CCBs [14-16]. Our study showed that this paradoxical sympathetic hyperactivity is caused by increment of neurotransmitter release from sympathetic neurons produced by L-type CCBs due to its interference on the Ca2+/cAMP interaction [5-8] (Figure 1).
In addition, several studies showed that increase of cytosolic cAMP concentration ([cAMP]c) stimulates neuroprotective response [17,18]. In this way, increase of [cAMP]c by interfering in the Ca2+/cAMP interaction could attenuate neuronal death triggered by cytosolic Ca2+ overload [5-8]. Then, the pharmacological handling of the Ca2+/cAMP interaction produced by combination of the L-type CCBs used in the antihypertensive therapy, and [cAMP]c enhancer compounds used in the anti-depressive therapy such as rolipram, could be a new pharmacological strategy for enhancing neurotransmission in neurological and psychiatric disorders resulting of neurotransmitter release deficit, and/or neuronal death [5-8]. These findings could open a new avenue for the drug development more effective and safer for the treatment of Alzheimer's and Parkinson's diseases.
Conclusion
Novel adventures throughout our discovery entitled "calcium paradox" due to Ca2+/cAMP interaction have been emerging to treat psychiatric and psychiatric disorders. Pharmacological handling of this interaction could be a more efficient and safer therapeutic strategy for stimulating neurotransmission compromised by neurotransmitter release deficit, and attenuating neuronal death [5-8,19-24].
Disclosure Statement
Caricati-Neto and Bergantin thank the continued financial support from CAPES, CNPq and FAPESP (Bergantin's Postdoctoral Fellowship FAPESP #2014/10274-3).
The authors also thank Elsevier - "author use": Reuse of portions or extracts from the article in other works - https://www.elsevier.com/__data/assets/pdf_file/0007/55654/AuthorUserRights.pdf.
References
- Douglas WW, Rubin RP (1961) The role of calcium in the secretory response of the adrenal medulla to acetylcholine. J Physiol 159: 40-57.
- Baker PF, Knight DE (1978) Calcium-dependent exocytosis in bovine adrenal medullary cells with leaky plasma membranes. Nature 276: 620-622.
- Neher E, Zucker RS (1993) Multiple calcium-dependent processes related to secretion in bovine chromaffin cells. Neuron 10: 21-30.
- Chern YJ, Kim KT, Slakey LL, et al. (1988) Adenosine receptors activate adenylate cyclase and enhance secretion from bovine adrenal chromaffin cells in the presence of forskolin. J Neurochem 50: 1484-1493.
- Caricati-Neto A, García AG, Bergantin LB (2015) Pharmacological implications of the Ca2+/cAMP signalling interaction: from risk for antihypertensive therapy to potential beneficial for neurological and psychiatric disorders. Pharmacol Res Perspect 3: e00181.
- Bergantin LB, Souza CF, Ferreira RM, et al. (2013) Novel model for "calcium paradox" in sympathetic transmission of smooth muscles: role of cyclic AMP pathway. Cell Calcium 54: 202-212.
- Bergantin LB, Jurkiewicz A, García AG, et al. (2015) A Calcium Paradox in the Context of Neurotransmission. Journal of Pharmacy and Pharmacology 3: 253-261.
- Bergantin LB, Caricati-Neto A (2016) Challenges for the pharmacological treatment of neurological and psychiatric disorders: Implications of the Ca2+/cAMP intracellular signalling interaction. Eur J Pharmacol 788: 255-260.
- Grossman E, Messerli FH (1998) Effect of calcium antagonists on sympathetic activity. Eur Heart J F: F27-F31.
- Kawamoto EM, Vivar C, Camandola S (2012) Physiology and pathology of calcium signaling in the brain. Front Pharmacol 3: 61.
- Wu CL, Wen SH (2016) A 10-year follow-up study of the association between calcium channel blocker use and the risk of dementia in elderly hypertensive patients. Medicine (Baltimore) 95: e4593.
- Hanon O, Pequignot R, Seux ML, et al. (2006) Relationship between antihypertensive drug therapy and cognitive function in elderly hypertensive patients with memory complaints. J Hypertens 24: 2101-2107.
- Forette F, Seux ML, Staessen JA, et al. (2002) The prevention of dementia with antihypertensive treatment: new evidence from the systolic hypertension in Europe (Syst-Eur) study. Arch Intern Med 162: 2046-2052.
- Kreye VA, Luth JB (1975) Proceedings: verapamil-induced phasic contractions of the isolated rat vas deferens. Naunyn Schmiedebergs Arch Pharmacol 287: R43.
- French AM, Scott NC (1981) A comparison of the effects of nifedipine and verapamil on rat vas deferens. Br J Pharmacol 73: 321-323.
- Moritoki H, Iwamoto T, Kanaya J, et al. (1987) Verapamil enhances the non-adrenergic twitch response of rat vas deferens. Eur J Pharmacol 140: 75-83.
- Sommer N, Loschmann PA, Northoff GH, et al. (1995) The antidepressant rolipram suppresses cytokine production and prevents autoimmune encephalomyelitis. Nat Med 1: 244-248.
- Xiao L, O'Callaghan JP, O'Donnell JM (2011) Effects of repeated treatment with phosphodiesterase-4 inhibitors on cAMP signaling, hippocampal cell proliferation, and behavior in the forced-swim test. J Pharmacol Exp Ther 338: 641-647.
- Bergantin LB, Caricati-Neto A (2016) Insight from "Calcium Paradox" due to Ca2+/cAMP Interaction: Novel Pharmacological Strategies for the Treatment of Depression. Int Arch Clin Pharmacol 2: 007.
- Bergantin LB, Caricati-Neto A (2016) Novel Insights for Therapy of Parkinson's disease: Pharmacological Modulation of the Ca2+/cAMP Signalling Interaction. Austin Neurol & Neurosci 1: 1009.
- Bergantin LB, Caricati-Neto A (2016) Recent advances in pharmacotherapy of neurological and psychiatric disorders promoted by discovery of the role of Ca2+/cAMP signaling interaction in the neurotransmission and neuroprotection. Adv Pharmac J 1: 66.
- Bergantin LB, Caricati-Neto A (2016) From discovering "calcium paradox" to Ca2+/cAMP interaction: Impact in human health and disease. Scholars' Press 108.
- Ritz B, Rhodes SL, Qian L, et al. (2010) L-type calcium channel blockers and Parkinson disease in Denmark. Ann Neurol 67: 600-606.
- Mosharov EV, Larsen KE, Kanter E, et al. (2009) Interplay between cytosolic dopamine, calcium, and alpha-synuclein causes selective death of substantia nigra neurons. Neuron 62: 218-229.
Corresponding Author
Leandro Bueno Bergantin, Laboratory of Autonomic and Cardiovascular Pharmacology, Department of Pharmacology, Escola Paulista de Medicina, Universidade Federal de São Paulo (UNIFESP), Rua Pedro de Toledo, 669 - Vila Clementino, São Paulo-SP, Brazil, Tel: 040-39-032, Fax: 55-11-5576-4973.
Copyright
© 2017 Bergantin LB. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.