Genetic and Molecular Analysis of the Genotypes of Bread Wheat and its Single Cross Hybrids
Abstract
A field experiment was conducted in the fields of a farmer on the right bank of the Euphrates River in the city of Ramadi/the center of Anbar Governorate, which is located at longitude 43° and latitude 33°. Five genotypes of bread wheat were used, it was introduced into diallel crosses in the winter season 2018-2019 to produce 10 single cross hybrids. Parents and their half-diallel hybrids were planted in the winter season 2019-2020 according to a randomized complete block design (RCBD) with three replications, with the aim of studying the effects of general and specific combining ability to some genetic parameters in addition to the ISSR molecular index. The variance of general and specific combining ability was significant in all studied traits. The G12 genotype was the best genotype in terms of general combining ability of grain yield, While the best hybrid in terms of special federation was (Babylon × Buhooth 10) in the yield of plant grains and (99 IPA × G12) in the number of grains per spike and the weight of 1000 grains. The ratio between the general combining ability variance (GCA) to the specific (SCA) was less than one for all the studied traits, this indicates that there is a dominant gene action that controls heredity. The values of dominance genetic variance were greater than the values of additive genetic variance for all studied traits, this was reflected in a decrease in the values of the heritability ratio in the narrow sense and an increase in the average degree of dominance over one for all the studied traits. With regard to the molecular analysis, the results show that the total produced bundles amounted to 93 total bundles, of which 85 were polymorphism, the molecular sizes of the replicated bundles ranged from 2200-350 bp. The two primers (UBC758 and UBC805) had the highest percentage of Polymorphism bundles of 100%. The new combinations can be used to produce single cross hybrids with a specific combining ability to produce a high yield, because most of her trait were under the influence of the dominance and over dominance, the ISSR index is highly efficient in diagnosing purity and genetic divergence between the genotypes of wheat.
Keywords
Polymorphism, Genetic, GCA, SCA, ISSR
Introduction
The main objective of most breeding programs for wheat is to increase the productivity of improved varieties, so it is important that the pure varieties and strains that are included in the formation of wheat crosses are distinguished by a high combining ability, in order to study its genetic behavior due to the presence of genetic differences between parents and their hybrids.
Wheat ( Triticum aestivum L) is the first crop in the world in terms of cultivated area and global production, despite the increase in the global rate of production of cereal crops such as wheat, barley and rice per unit area for human nutrition to nearly double what it was at the beginning of the twentieth century, as a result of the efforts of researchers and those interested in breeding and crops improving.
However, the gap between global production and demand still exists and is constantly increasing, especially if we know that the world population has reached more than seven billion people by the beginning of the year 2020, and that two-thirds of the increase in population is in the third world countries [1-3]. Success in solving agricultural problems is mainly related to an increase in production. This increase is achieved through the expansion of the cultivated area and the use of modern farming methods on the one hand, and the use of new varieties with high productivity and resistance to biotic and a biotic stresses on the other hand. The main objective in most breeding programs is to improve the genetic potential of the grain yield and its components. The first stages of breeding are the most difficult, as the plant breeder faces many difficulties in determining the parental phenotypes that will enter the process of crossbreeding and selecting the best genotypes in the segregation generations of the hybrid groups [4]. The trait of grain yield is a complex quantitative trait whose heredity is controlled by a large number of genetic factors, so it is necessary to know which type of gene action most contribute to improvement. This is done by knowing the components of genetic variance for different traits. To reach the goal, breeding programs are designed that depend on crossbreeding and on the ability to compatibility between the parents involved in crossbreeding and knowing the possibility of achieving genetic gain in a short time [5].
Half-Diallel Crosses method between selected cultivars is one of the methods used in collecting more than one trait to obtain a superior hybrid. It is one of the important methods in analyzing genetic experiments that are used for the purpose of identifying the genetic behavior of the studied traits of parents and hybrids within the genetic improvement program [6].
Many researchers have carried out diallel crosses and found there were significant differences in the effects of general and specific combining ability in the traits they studied [7-10].
The efforts of plant breeders continue to develop and increase wheat productivity and genetic improvement by using modern biological techniques based on genotypes DNA, studying molecular indicators and developing breeding and improvement programs. However, there is little information available about the wheat genome because of its large size in wheat, which is estimated at 1.7 × 1010 pb [11]. Molecular index is one of the important indexes, depending on the ISSR (Inter Simple Sequence Repeats) technology, which can determine genetic relationship, measure genetic diversity, and detect genetic differences between types and varieties of field crops, and conduct hybridization between genotypes based on their results, which can be trusted. It gives a quick visualization of the genome sequence and then reveals the genetic differences that individuals carry [12]. It is one of the important studies in the programs of breeding and crops improvement for the purpose of selecting the most appropriate parents to obtain the appropriate hybrids, in addition to describing the genetic material and molecular parameters.
This study aims to estimate the general and specific combing ability, genetic variance, heritability, and the degree of dominance of the traits of the yield and its components in bread wheat in order to benefit later in the breeding programs when selecting individuals with desirable traits that can be developed into commercial varieties and molecular characterization at the DNA level using ISSR technology.
Materials and Methods
Five wheat genotypes were used in this study, numbers were given from 1 to 5, entered into half-diallel cross-breeding. They are (1) IPA 99 (2) Bohooth 10 (3) Babel (4) G5 (5) G12 (Table 1). Parents seeds were sown on lines in a farmer’s fields next to the Euphrates River, located at longitude 43° and latitude 33°, in the winter season 2018-2019, with six lines for each genotype, with a line length of 4m. The seeds were sown in hills at a distance of 15 cm and between a line and another 60 cm. Crossing between parents was carried out with all possibilities in one direction, at the end of the season 10 single crosses were obtained by the method described by Griffing [6] by the half-diallel cross method.
In the winter season 2019-2020, parent’s seeds and hybrids were planted in the same area according to a randomized complete block design (RCBD) with three replications, each replicate contained five parents and ten single crosses. The experimental unit included 4 lines, and the following traits were studied on the basis of one plant for the ten random plants: The area of the flag leaf and the number of spikes m -2 , the number of grains in the spike, the weight of 1000 grains, and the grain yield of the plant, the data obtained from the parents and their individual crosses of the studied traits were analyzed according to the design used. Then the parents' and hybrids data were analyzed according to the first model of the second method, and the following equation was used P(P+1) /2 (where P = number of parents), to determine the number of genotypes included in the experimental units and that come up with 15 genotype in each experimental unit.
The sum of the squares of the general combining ability was estimated, as well as the estimation of the sum of the squares of the specific combining ability. The values of the additive variance б 2 A, the dominance б 2 D and the environmental б 2 e were estimated as follows: [13]
б 2 e = Mse, б 2 D = б 2 sca, б 2 A = 2б 2 gca
The values of heritability in the broad and narrow sense and the average degree of dominance were also estimated by means of the expected mean variance from the Griffing analysis.
DNA extraction
The process of DNA extraction from young leaves was carried out using (CTAB) substance according to what was mentioned [11,14].
ISSR index (inter simple sequence repeats)
The basics of searching for variation at the level of DNA replicated by PCR reaction depend on the concepts of the appearance of bands in DNA index in general and ISSR index in particular. The most important of which is the absence of results for doubling in the PCR reaction, the appearance of results of equal sizes and numbers, the appearance of results of different sizes and different numbers, and the appearance of results with packages of different densities. Eight primers were used (Table 2).
Results and Discussion
The existence of significant differences between parents and their crosses makes it possible to continue studying the genetic behavior of them. The results of the analysis of variance indicate that there are significant differences for all the studied traits, which indicates the presence of genetic differences between the parents, which requires the evaluation of these parents (Table 3). It also appears from the results of the analysis of the general and specific combining ability that the differences between the mean squares of the general and special combining ability were significant for all the studied traits, this explains the importance of both additive and non-additive effects on the studied traits. The variance of the general combining ability to the variance of the specific combining ability (б 2 gca / б 2 sca) was less than one true for all studied traits, which indicates that the non-additive effects were more important than the additive effects. It can be inferred that there is non-additive gene action that controls the inheritance of the studied traits. These results are consistent with what others have found [7,12,15-17].
These differences reflect the number of genetic differences, which requires evaluating the parents and the resulting hybrids (Table 4). The studies conducted confirmed the existence of significant differences between individual crosses and their parents for several studied traits [18].
The parent (1 and 4) had the highest value among the parents in the area of the flag leaf, and the two parents (3 and 5) were superior in the number of spikes m -2 . The parents (4 and 5) were superior in the number of grains in the spike, while the parents (1 and 2) outperformed in the weight of 1000 grains, in the grain yield of the plant, the two parents (1 and 5) were superior by giving them the highest grain yield per plant, which reached 36.74 and 36.70 gm, respectively. Whereas for the hybrids, the hybrid (3 × 2) gave the widest mean of the area of the flag leaf (97.8 cm 2 ) and did not differ significantly with two hybrids, while the hybrid (5 × 1) showed the highest number of spikes m -2 (360.4) and did not differ significantly with four crosses as well, the same hybrid gave the highest weight of 1000 grains (48.9g). As for the number of grains in the spike, the hybrid (4 × 2) was distinguished by the highest number of grains in the spike (69.7), while the hybrid (5 × 1) was distinguished by the highest yield per plant (47.36g). This hybrid was distinguished by the high yield, perhaps because of its superiority in the number of spikes m -2 and the weight of 1000 grains. This hybrid did not differ significantly with the hybrids (5 × 4) and (3 × 2). These results agree with the findings of several researchers [19,20] as this is due to the genetic divergence between the parents involved in the hybrid.
The estimates of the effect of the general combining ability of parents in the traits studied. The parent (5) gave a positive and significant effect, and was the best parent in the grain yield of the plant (1.58), the area of the flag leaf is (2.37) and the weight of 1000 grains is (1.15). While parent (1) was the best in the number of spikes m -2 (7.90), while parent (4) gave the highest positive and significant effect on the number of grains per spike (2.18) (Table 5). It is possible to benefit from the best parents because they possess the desired genes and contribute significantly to the transfer of the trait to their hybrids. These results are consistent with what was found [3,9,21] that the general combining ability has significant effects in some parents in transferring some of the studied traits.
When calculating the effects of the specific combining ability of hybrids, where the hybrid (3 × 2) was the best in the area of the flag leaf (4.87), the number of spikes m -2 (29.41) and the plant yield (9.33). As for the hybrid (5 × 1), it was the best in the number of grains per spike (3.58) and the weight of 1000 grains (7.86). Some of the parents gave high values for the specific combining ability for some traits when crossed with other parents (Table 6).
This is consistent with the findings of others [15,22] that some hybrids show a positive and significant influence, while others show a negative and significant influence on the specific combining ability in the studied traits. Parents that had a positive and significant effect on the general combining ability to a trait gave significant effects in the same direction in the effects of their hybrids for specific combining ability, meaning the occurrence of the dominance effect of genes. But if the effects of general combining ability are positive and significant for a trait and no positive effect of specific combining ability is produced, then this is caused by the effect of the additive gene action of those traits. If the specific combining ability is more important than the general combining ability, then this means that genetic control is for the dominance gene action.
When estimating the values of the components of phenotypic variance (σ 2 P), it was noted that the values of the additive genetic variance σ 2 A, the dominant σ 2 D and the environmental variance σ 2 e differed from zero for all traits. Values of the dominance genetic variance were higher than their additive counterpart for all traits, which indicates that the genes related to the dominant genetic variance were more important than the genes related to the additive genetic variance in controlling the inheritance of traits characterized by high dominance genetic variance (Table 7). This result is in agreement with the other findings [15,10,22,23].
The percentage of heritability in its broad sense was high for all traits, as it ranged between 89.28% for the number of grains per spike and 96.83% for the area of the flag leaf, this is due to the high values of genetic variance and low environmental variance values, which makes the possibility of passing on the trait to individuals of the first generation more likely. The values of heritability in the narrow sense were moderately high for the number of spikes m -2 (12.31%), the number of grains per spike (12.29%) and the grain yield (12.68%), and the reason for this is due to the high values of additive genetic variance, so it can be improved by selection, whereas for the rest of the traits, the percentage of heritability in the narrow sense ranged between (3.23%) for the area of the flag leaf and (3.89%) for the weight of 1000 grains, so it can be improved by hybridization. As for the average degree of dominance, it was greater than one for all the traits studied, and this indicates the presence of the effects of the over-dominance of the genes that control the inheritance of these traits [4,17].
ISSR primer results
Results of the eight primers run on agarose gel that the primers included in the study were produced the total number of bundles amounted to (93) total bundles, of which (85) were Polymorphism from the total bundles. The primers (UBC686 and UBC758) produced the highest number of bundles, reaching (13) bundle, while the primer (A41) produced the least number of bundles, reaching (10) bundles. The primers UBC758 (and UBC805) produced the highest percentage of Polymorphism bundles (100%). The lowest is in the primer (A41) as it reached (80%), the greater the number of Polymorphism bundles.
The lowest was in the initiator (A41) as it reached (80%). The greater the number of polymorphism bundles, the greater the efficiency of the primers in determining the genetic diverge and the possibility of obtaining the genetic fingerprint of the genotypes included in the study. This agrees with [24-26] as the total of the absent bundles was (14) bundles, the primer (UBC686) had the most primer in the number of absent bundles, which amounted to (4) absent bundles. The number of unique bundles is (3) bundles, which is a distinguishing feature for the varieties, and their appearance is an indication of the occurrence of mutations and new unions that produce hybrids in the plant gene, which determines the genetic fingerprint of the variety [26], variation in bundles is the result of induced mutations or self-mutations that affect the distance between genetic loci that occur naturally during the evolutionary process that occurs in living organisms, the variation may have occurred by addition or deletion at the sites to which the primer is attached, This is what found in other studies [27,28], and the molecular sizes of the multiplicative bundles of the primers included in the study ranged between (2200-350 bp). This agrees with [28] (Table 8).
Genetic diverge based on molecular markers
The genetic dimension was estimated using genetic program NTSYS-PC version 2.1, which depends on the amount of similarity or difference between the genotypes. It was found through the values of the genetic diversity referred to in Table 9 the values of the genetic diversity of the genotypes included in the study when eight primers were used randomly. If the genetic material is identical between two genotypes, this indicates that the genetic diversity between them must be equal to zero. As for the percentage of genetic similarity, it represents the measure of the degree of genetic similarity, therefore, they are equal when there is no genetic variance between the genotypes, and this appears when individuals participate in the general bands and no polymorphism bands appear between them. What diminishes the interest in such findings is the use of so few primers, and when using more than one primer, due to the different regions of association according to the primer sequence. What determines the genetic similarity or genetic diversity between genotypes is the number of shared bundles.
The greater the number of those bundles, the lower the genetic diversity and viceversa. Those bundles that share with each other indicate the similarity of the genetic material in the genome region to the structures included in the study, which may represent the similarity in phenotypic traits. Or the similarity may be in the non-coding regions, that is, those that do not have gene expression. Whereas for the genetically divergent genotypes, they are the ones that share the least number of bundles with each other due to differences in the nucleotide sequences in the genome [20], the values of the genetic diversity ranged between (0.115-0. 775), where the lowest genetic diversity was between the two genotypes (Babel and IPA 99), where it reached (0.115), this is the highest similarity between the two genotypes included in the study. The largest genetic diversity reached (0.775) between the two genotypes (G12 and 99 parents), and this is the least genetic similarity between the two genotypes, while the values of the genetic diversity for the rest of the genotypes ranged between those values as shown in Table 9.
Cluster and of principal components analysis based on molecular markers
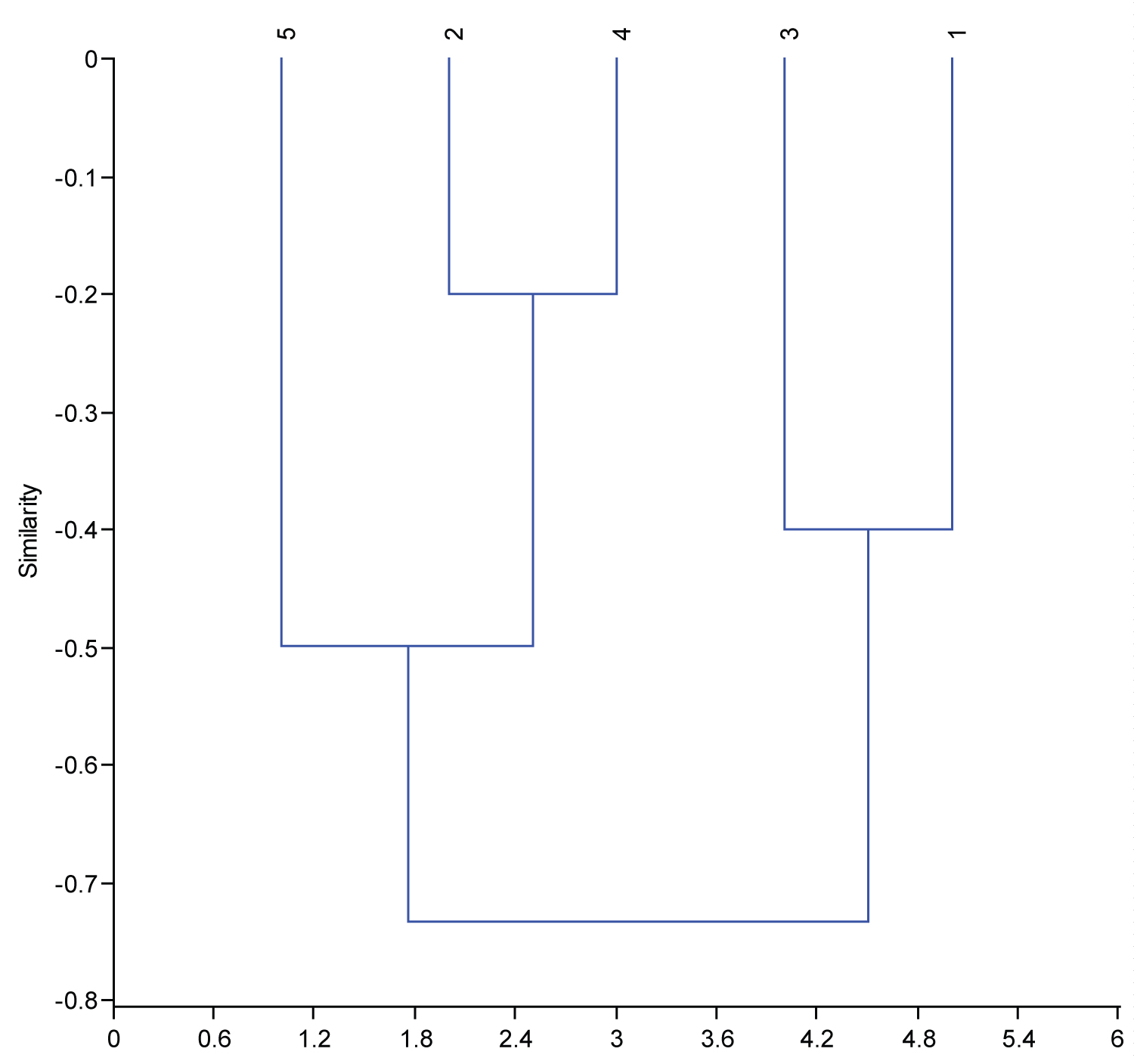
The order of the genotypes, depending on the values of the genetic diversity, the dendrogram, depends on the genetic range in which the main groups are linked. Therefore, the presence of group of the genotypes in a particular group indicates the extent of the genetic similarity to those structures in that group [26].
The cluster diagram shown in Figure 1 represents the five genotypes of the bread wheat crop used in the study. It was divided into two main groups, the first group included the two genotypes (IPA 99 and Babel), while the second main group was divided into two secondary groups, as the first secondary group included the two genotypes (Buhooth 10 and G5), while the second secondary group was divided into the genotype group (G12).
Through these results, it was found that there is a degree of genetic similarity or diversity between the genotypes included in the study, and this is due to two things, first the difference between the five genotypes may be in the phenotypes, and this is the result of the influence of the environment, which is what depends on it in many breeding programs, second it may be the result of a genetically high similarity between the genotypes, which contradicts the first thing, which depends on phenotypes, and this is due to the similarity in the non-coding regions of genes [27]. This is consistent with many researchers who used the indicators of molecular markers, especially the ISSR, including the study [12,24,26,28,29].
Conclusion
It can be concluded from the above that the parent G12 (5) is the best of the parents because its general combining ability was high for the flag leaf area, weight of 1000 grains and the yield of individual plant, and that the hybrid (IPA99 × G12) is the best among the hybrids for giving it a high grain yield for individual plant (47.36 gm), its specific combining ability was high for a number of traits, and the superiority of this hybrid in grain yield is due to its superiority in the area of the flag leaf, the number of spikes m -2 , and the weight of 1000 grains. We conclude that the molecular study and the ISSR index showed that the genotypes of wheat contain a high genetic diversity, and these results can be used as a basic material in breeding programs using hybridization to improve the quantitative and qualitative traits.
Acknowledgement
We would like to extend our thanks to everyone who contributed to the completion of this research. Our special thanks to the owner of the private farm who provided us with all the available conditions. Thanks, and appreciation to those who supplied us with the wheat varieties under study. All appreciation goes to the Center for Desert Studies and its staff for assigning this work.
References
- Abas SA, Abood NM, Al-Hamed ZA, et al. (2018) Heterosis, combining ability and some genetic parameters in wheat using half diallel mating design. J Plant Production 9: 1-5.
- Abd HS, Abdulhamed ZA, Ghadir MA, et al. (2021) Estimation of genetic parameter by using full diallel cross in maize under different irrigation interval. IOP Conference Series: Earth Environ Sci 904: 012001.
- Abdulhamed ZA, Abdulkareem BM, Noaman AH, et al. (2021) Efficiency of ISSR markers detect geneic and molecular variation between barley genotypes. Int J Agric Stat Sci 17: 1503-1508.
- Abdulhamed ZA, Alfalahi AO, Abood NM, et al. (2020) Riboflavin and cultivars affecting genetic parameters in maize (Zea mays L.) AIP Conference Proceedings 2290: 020020.
- Verma AK, Mehta AK, Singh RP, et al. (2020) Studies on hybrid vigour for yield and contributing traits in cowpea (Vigna unguiculata L. Walp). Res J Biotechnol 15: 223-231.
- Griffing B (1956) Concept of general and specific combining ability in relation to diallel crossing systems. Aust J Bio Sci 9: 463-493.
- Abebe AA, Desta TM (2017) Genetic variability and character association of some local wheat varieties (Triticum species L.) using agro morphological traits grown in South Gondar zone, Ethiopia. African. J Biotechnol 16: 2100-2105.
- Fellahi ZE, Hannachi AA, Bouzerzour H, et al. (2018) Analysis of direct and indirect selection and indices in bread wheat (Triticum aestivum L.) segregating progeny. Int J Agro 1: 1-12.
- Abdulhamed ZA, Abas SA, Abed AA, et al. (2020) Studying some genetic in maize by line x tester analysis. Int Agricult Stat Sci Supplement 1: 1421-1426.
- Abdulhamed ZA, Abood NM (2021) Combining ability and gene action in sunflower using cytoplasmic male sterility. Indian J Ecol 15: 223-228.
- Huang QX, Wang XC, Kong H, et al. (2013) An efficient DNA isolation method for tropical plants. Afr J Biotechnol 12: 2727-2732.
- Farhan MB, Abdulhamed ZA, Noaman AH, et al. (2019) Determination of genetic distance among genotypes of bread wheat Triticum aestivum L using issr markers. J Plant Archives 19: 455-459.
- Singh RK, Chaudhary BD (2010) Biometrical Methods in Quantitative Genetic Analysis. Kalyani Publishers, New Delhi - Ludhiana, India, 318.
- Weigand F, Baum M, Udupa S, et al. (1993) DNA molecular marker techniques, technical manual. No.20. International Center for Agricultural Research in the Dry Area (ICARDA). Aleppo Syria.
- Al-Kaabi DHH (2015) Characterization of some Wheat Varieties Biotechnology. Ph. D. Dissertation. Coll. Of. Agric. Univ of Baghdad 164.
- Grzesiak S, Hordynska N, Szezyrek P, et al. (2019) Variation among wheat (Triticum aesativum L.) genotypes in response to the drought stress: I-selection approaches. J Plant Interac 14: 30-44.
- Ly D, Huet S, Gauffreteau A, et al. (2018) Whole-genome prediction of reaction norms to environmental stress in bread wheat (Triticum aestivum) by genomic random regression. Field Crops Res 216: 32-41.
- Dashchi S, Abdollahhi BM, Darvishzade R, et al. (2016) Molecular similarity relationships among Iranian bread wheat cultivars and breeding lines using ISSR markers. Not Bot Horti Agrobo 50: 254-260.
- Kumar P, Singh G, Singh YP, et al. (2015) Study of combining ability analysis in half diallel crosses of spring wheat (Triticum aestivum L.) Int J Adv Res 3: 1363-1370.
- Al-Assaf, Latif A, Hakim MS, et al. (2019) Study of genetic variances and trait correlations for genotypes of barley (Hordeum vulgare L.) under the conditions of wet and dry environments in Syria. Syrian J Agric Res 6: 254-269.
- Amoon MH, Abdulhamed ZA (2000) Determination genetic diversity of inbred lines and hybrids of maize using issr technic. Iraqi J Agri Sci 51: 269-277.
- Mohamed AMA, Abdulhamed ZA, et al. (2021) RAPD to estimate genetic distance in some maize genotypes. Indian J Ecol 15: 255-260.
- Abdulhamed ZA, Abood NM, Noaman AH, et al. (2021) Genetic analysis and correlation studies of yield and its components of some bread wheat varieties. IOP Conf Seri: Earth Environ Sci 761: 012066.
- Khaled AGA, Motawea MH, AA Said, et al. (2015) Identification of ISSR and RAPD markers linked to yield traits in bread wheat under normal and drought conditions. J Genet Eng Biotechnol 13: 243-252.
- Al-timime AHT (2019) Genetic Diversity by RAPD Technique, Line x Tester and Somatic Hybridization in bread wheat. PhD. Dissertation Coll. Of Agric. Univ. of Baghdad 199.
- Reham, A AL-K, Ghinwa L, Al-Husien N (2019) Genetic diversity of some wheat (triticum durum) genotypes using SSR technique. J Syrian Agric Res 6: 68-83.
- Alsaman B, Wjhani Y, Ch Albasatneh M, et al. (2015) Evaluation of some atriplex accessions under salt stress conditions and their molecular characterization using ISSR technique. Syrian J Agric Res 2: 22-31.
- Majeed DM, Ismail EN, Al-Mishhadani I I, et al. (2018) Assessment of genetic diversity among wheat selected genotypes and local varieties for salt tolerance by using RAPD and ISSR analysis. Iraqi J Sci 59: 278-286.
- Abdulhamed ZA, Abas SA, Noaman AH, et al. (2021) Genetic Performance of Inbred and Hybrids of Maize under Irrigation Interval. IOP Conf Ser Earth Environ Sci 904: 012001.
Corresponding Author
Ghazwan Husam Tawfeeq, Department of Soil Sciences and Water Resources, College of Agriculture, University of Anbar, Ramadi, Iraq.
Copyright
© 2023 Abdulhamed ZA, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.