Himalayan Pink Salt as a Healthier Substitute in Lowering the Risk of Dietary Salt Induced Cognitive Insults in Experimental Rats
Abstract
Alzheimer's disease (AD) belonging to the class of tauopathies, is a major cause of dementia globally. Observational studies in the aging population have reported that excessive dietary salt consumption is one of the risk factors that contribute to the pathological processes that underlie dementia and cognitive impairment. Regulation of the daily salt intake and consumption of alternative nutritional salts, if proven, may serve as a cost-effective and feasible approach in lowering the risk of dementia. With this background, the objective of the current study included assessment of the putative benefits of Himalayan pink salt with regard to its effects on spatial memory in comparison with that of common salt using rat as an experimental model. The results of the study showed that Himalayan pink salt, even at 5-10 fold higher concentrations of the salt over the permissible daily intake limits, could exhibit a positive effect on learning and memory in Barnes maze task and improved nitric oxide levels. Phosphorylated tau levels remained comparable to that of control group whereas common salt inflicted greater insults leading to elevated levels of pathological form of tau and compromised spatial memory. These findings suggest that Himalayan pink salt may serve as a healthier salt substitute towards lowering the risk or severity of dietary salt induced form of AD.
Keywords
Himalayan pink salt, Table salt, Spatial memory, Phospho-tau, Salt inducible kinase, Sporadic Alzheimer disease
Introduction
The salt consumption in many countries worldwide exceeds the recommended daily intake levels. In Asia-Pacific region the daily salt intake ranges between 9.1-11.0 g/day in countries such as Japan, Australia, and China. The mean population salt consumption in India spans 9.45-10.41 g/day [1]. The current daily global salt consumption is almost double the upper limit of < 5.0 g/day that is recommended as per the World Health Organisation (WHO) guidelines [2].
Hypertension has long been associated with excessive salt consumption [3]. Chronic hypertension is a vascular risk factor for stroke, cerebrovascular diseases, Alzheimer's disease (AD) and other tauopathies [4]. As in case of tauopathies and AD, accumulation of hyper phosphorylated tau is also observed in cerebrovascular disorders associated with endothelial dysfunction and cognitive impairment [5]. The pathology of AD is characterized by the presence of extracellular amyloid beta plaques, neurofibrillary tangles containing hyper phosphorylated tau, and neuronal cell death [6]. Familial AD is a rare, early-onset form which has been established to have strong links to mutations of presenilin (PSEN 1 & 2) and amyloid precursor protein (APP) genes. In contrast, the sporadic form of AD is late-onset and multifactorial. It is associated with diet, hypertension, metabolic syndrome, infections, mitochondrial dysfunction, immune system, and vascular disease, allelic variations of Apo E gene, traumatic brain injury, metal exposure and poor quality of sleep [7].
As per the recommendation of WHO, salt reduction is one of the most feasible and cost-effective approaches for the prevention of non-communicable diseases (NCDs) [8] including AD. In this light, low sodium salts as well as alternative salt varieties such as the Himalayan pink salt [9] are being promoted as nutritionally superior substitutes to common salt [10]. These salts have a number of minerals such as magnesium, potassium and calcium in addition to sodium and chloride [11].
In the current study, the putative health benefits of the acclaimed, nutritionally superior mineral salt i.e., Himalayan pink salt on spatial learning and memory was investigated. High concentration of common salt which is known to contribute to endothelial and neuronal dysfunction was also included in this study. Five to ten-fold higher concentrations of the salt over the permissible daily intake limits were orally administered per day to 4-month-old female SD rats for 3 months. Thereafter, the memory impairments were assessed by Barnes maze task. The serum nitric oxide metabolites as a measure of endothelial dysfunction were estimated. High salt induced alterations in the kinases likely involved in tau hyperphosphorylation were investigated. The results of these assessments are in favour of Himalayan pink salt as a healthier salt substitute towards lowering the risk or severity of AD.
Materials and Methods
Chemicals
Himalayan pink salt (Nutroactive brand) and common table salt were sourced through e-commerce platform. Nitrite/nitrate assay kit, phenylmethylsulfonyl fluoride (PMSF) were purchased from Sigma Aldrich, Bengaluru. Rat Serine/Threonine-Protein Kinase SIK1 (SIK1) ELISA Kit, Rat Phospho Tau Protein ELISA Kit and Rat Tau Protein (TAU) Elisa kit were procured from MyBioSource, Inc., USA. Other reagents used were of analytical grade and obtained locally.
Animals
4-Month-old Sprague Dawley (SD) female rats weighing 200-300g (n = 20) were procured from Central Animal Research Facility in the Institute. All the experiments involving animal usage was approved by the Institute animal ethics committee (IAEC number - AEC/71/464/NC). All the animals were housed in the standard animal facility with controlled environment of light-dark (12h:12h) schedule and had free access to pellet food and water ad libitum.
Chronic treatment of rats with high salt
1 mL volumes of freshly prepared salt solutions at two different supra-physiological concentrations were administered to the experimental groups. The salt solution was orally administered using an oral gavage tube. The control rats did not receive any treatment (Control)) and the experimental rats received salt concentrations of either 0.2 g/Kg body weight per day of Himalayan pink salt (PS 0.2) or 1.0 g/ Kg body weight per day of Himalayan pink salt (PS 1.0) or common salt (CS 0.2 or CS 1.0 respectively) for a duration of 12 weeks (n = 4 for each group).
Sample collection
The animals were anesthetized in a glass chamber using 0.5 mL of inhalant anaesthesia isoflurane, and then 1 mL of blood was obtained by cardiac puncture. The serum was separated by centrifugation at 4 ℃ for 15 min at 5000 rpm. After dissection, hippocampal tissue was homogenized with the buffer (25 mM Tris, pH 7.4 containing 20 µM EDTA, 3.25 mM NaCl, 100 µM PMSF). Clarified serum and hippocampal homogenate were subjected to biochemical assays.
Biochemical assays
The levels of nitric oxide (NO) metabolites in serum were measured using the Nitrite/Nitrate assay kit (sensitivity range: 10-100 µM) as per the manufacturer's instructions. The level of salt inducible kinase-1 (SIK-1) in the rat hippocampal homogenate was measured using the quantitative sandwich ELISA kit (detection range: 0.5 ng - 16 ng/mL). The levels of phospho tau protein in the serum were measured using the Rat Phospho Tau protein ELISA kit (detection range: 1.0 ng - 400 ng/ mL). Total tau protein in the rat serum was measured using the Rat Tau protein ELISA kit (detection range: 1.0 ng – 25 ng/mL) as per the manufacturer's instructions.
Spatial memory assessment
Hippocampal dependent spatial memory assessment was performed by subjecting animals to Barnes maze task as described by Joseph, et al. [12]. The maze consisted of an elevated circular platform of 120 cm in diameter with twelve equidistant holes along the circumference. Two 150W lights were hung above the platform to create an aversive environment which encourages the rats to escape from the brightly lit open platform to the dark environment of the escape box. The location of the escape box remained constant for any individual rat across the test trials during acquisition and retention. On day 1 of the behavioural assessment, experimental animals were first habituated to Barnes maze by placing them individually on the circular platform for 2 min. Day 2-4 included acquisition training consisting of a total of 15 trials (3 trials/day) and learning was assessed on every day. Post the completion of acquisition trials for 3 days, testing was abated for 4 days after which retention of memory was evaluated in three trials (1 day) and the mean average performance of the three trials was calculated. Latency (time taken by the rats to find the escape hole) and total errors (nose pokes into non-escape holes as well as nose pokes into escape hole) were recorded. In the case of experimental rat being unable to locate the escape box within 2 mins, it was gently guided to the target box. After 30 sec, the rat was removed from the escape box and returned to its home cage. After every trial, the platform and escape box were cleaned with 70% ethanol to eliminate olfactory cues left behind by the animal during the experiment.
Statistical analysis
All data were analysed with Graph Pad prism 8.0. Results of each of the biochemical analyses i.e., assessment of nitric oxide metabolites (NO) in serum, assessment of SIK1 levels in hippocampal tissue, and assessment of phospho-Tau level in serum between two groups were analysed by unpaired t test. Results of acquisition and recall test trials of Barnes maze task were analysed by one way ANOVA followed by Tukey's multiples comparison post hoc test. P values < 0.05 were considered statistically significant.
Results
Mineral composition of Himalayan pink salt that has been selected for the current study is presented in Table 1. It is richer in calcium, potassium and iron contents with marginally lesser sodium levels in comparison to common salt.
Assessment of nitric oxide (NO) metabolites in the serum
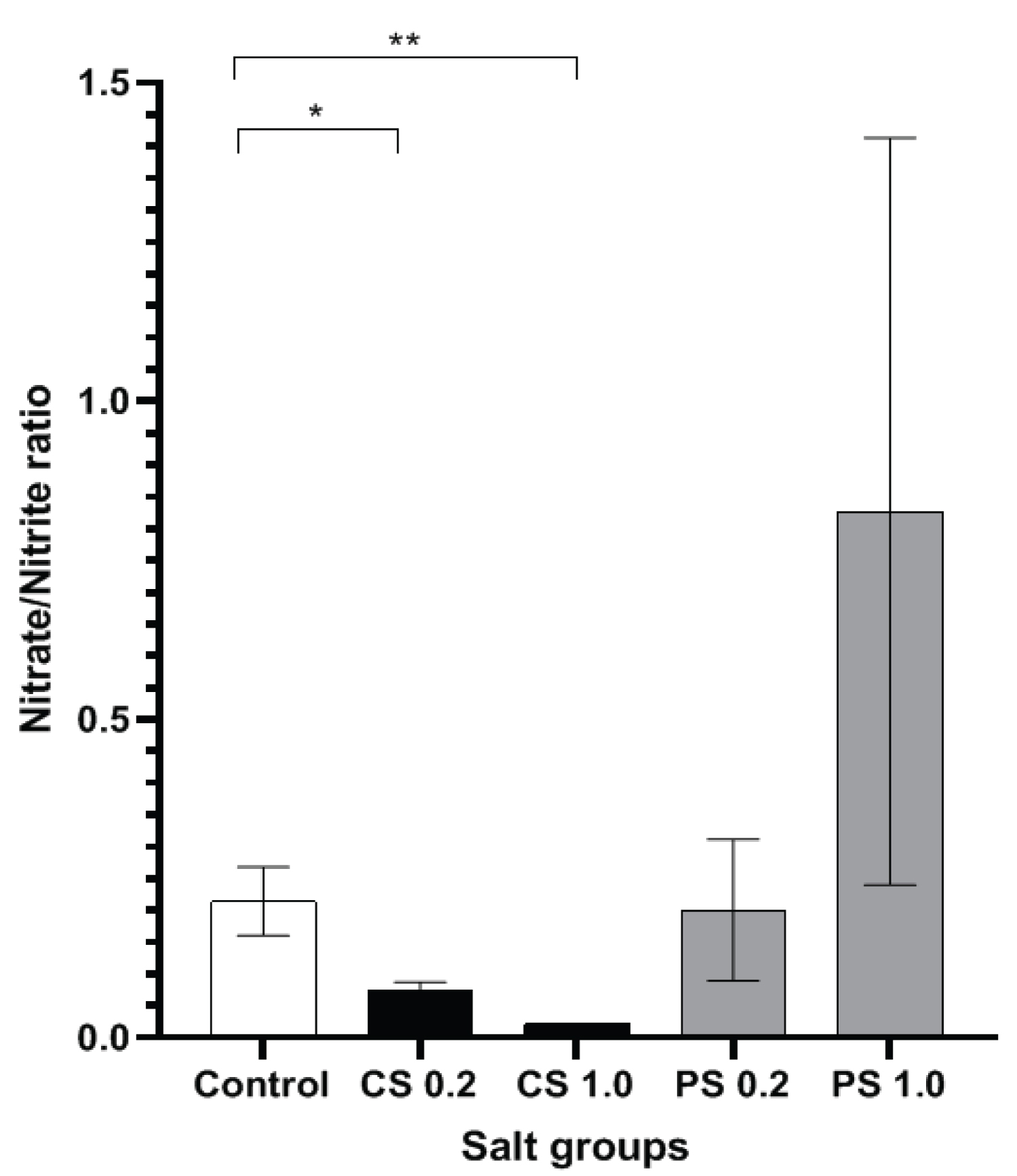
In the current study, to evaluate the high salt mediated endothelial dysfunction, serum NO metabolite levels were measured, and the ratio of nitrate/nitrite levels were assessed for each of the groups. Both 1.0 g/Kg body weight and 0.2 g/Kg body weight treated common salt groups had significantly lower serum nitrate/nitrite ratio compared to the control group with a more pronounced reduction shown by the 1.0 g/Kg body weight common salt group (Figure 1).
Assessment of spatial memory using Barnes maze task
Barnes maze is a dry-land behavioural test developed by Carol Barnes in 1979 to assess spatial memory in rats [13]. In this procedure the hippocampal-dependent spatial reference memory of the rats is assessed by their ability to recall the relationship between distal cues in the surrounding environment and the location of a fixed escape hole.
The different high salt experimental groups were evaluated by the Barnes maze test for spatial memory acquisition and retention at the end of 3 months of chronic high salt administration. The performance of each experimental group in the maze task was assessed using the number of errors (nose pokes into the non-escape holes) and the latency (the time taken to locate the escape box) during the acquisition (learning) phase and retention (probe) test and compared with the performance of the control group.
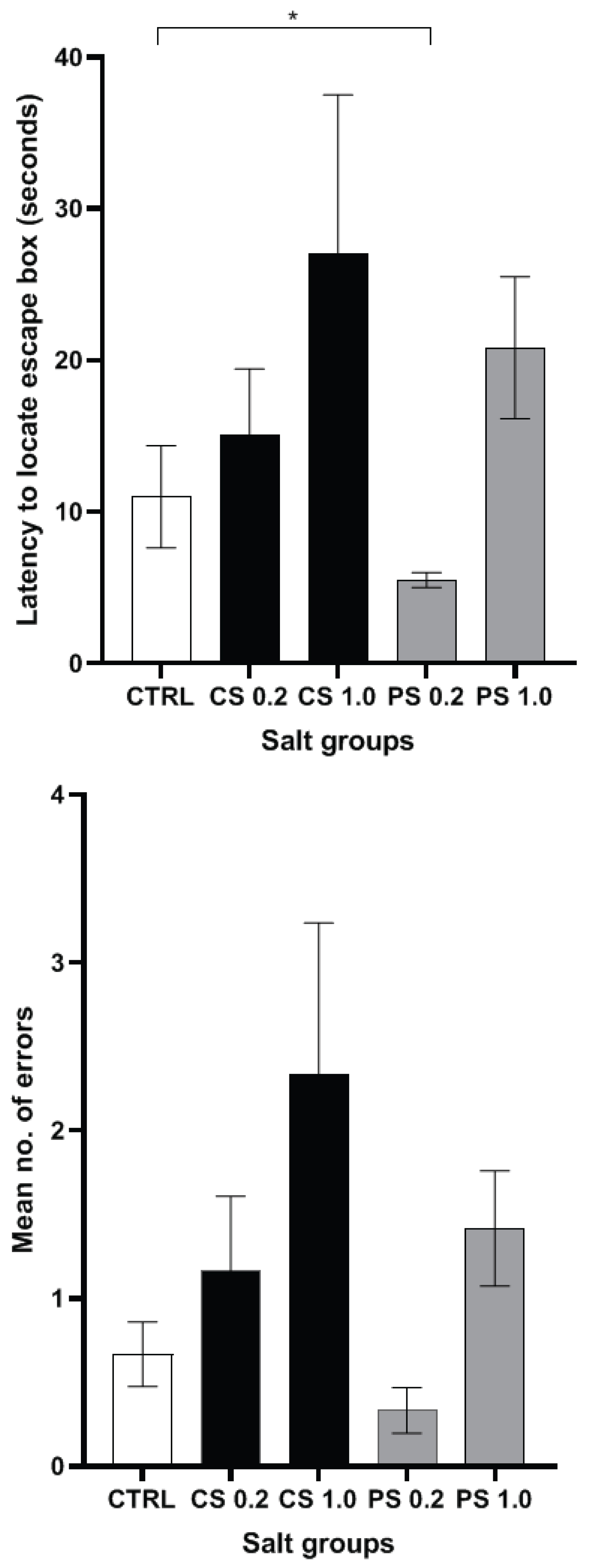
During the acquisition phase, from day 1 to day 3 of the trials (Table 2), as the training progressed, all the animals showed consistant and statistically significant improvement in learning the task, as reflected by reduced latency by Day 3. It is noteworthy that, in spite of chronic consumption of 0.2 g/Kg body weight/day of Himalayan pink salt, these animals were faster that untreated (no salt) control group in quickly learning to find the escape box, as demonstrated by significantly decreased latency as well as lesser number of errors committed in locating the escape box on Day 1 of acquisition trials (Table 3).
However, at the higher concentration of 1.0 g/Kg, the Himalayan pink salt group had higher latencies coupled with higher mean number of errors compared to the control group on each trial day although there was improvement in their respective learning parameters from day 1 to day 3. On the day of the recall test (Figure 2A and Figure 2B), although higher concentration of 1.0 g/Kg treated Himalayan pink salt group showed greater latency and greater number of errors similar to the common salt group, the results were statistically not significant, when compared to untreated control group.
Importantly, during the recall phase, despite consuming additional amount of Himalayan pink salt at a concentration of 0.2 g/Kg body weight per day for a duration of 12 weeks, this group had shown remarkable reduction in latency (p = 0.0173) (Figure 2A). It is noted that, although statistically not significant, pink salt group (0.2 g/Kg body weight) committed lesser number of errors (Figure 2B) compared to the control group and other treatment groups. These findings strengthen the case for considering pink salt as a superior alternative to common salt for spatial memory functions.
Measurement of salt inducible kinase 1(SIK1) levels in hippocampal tissue
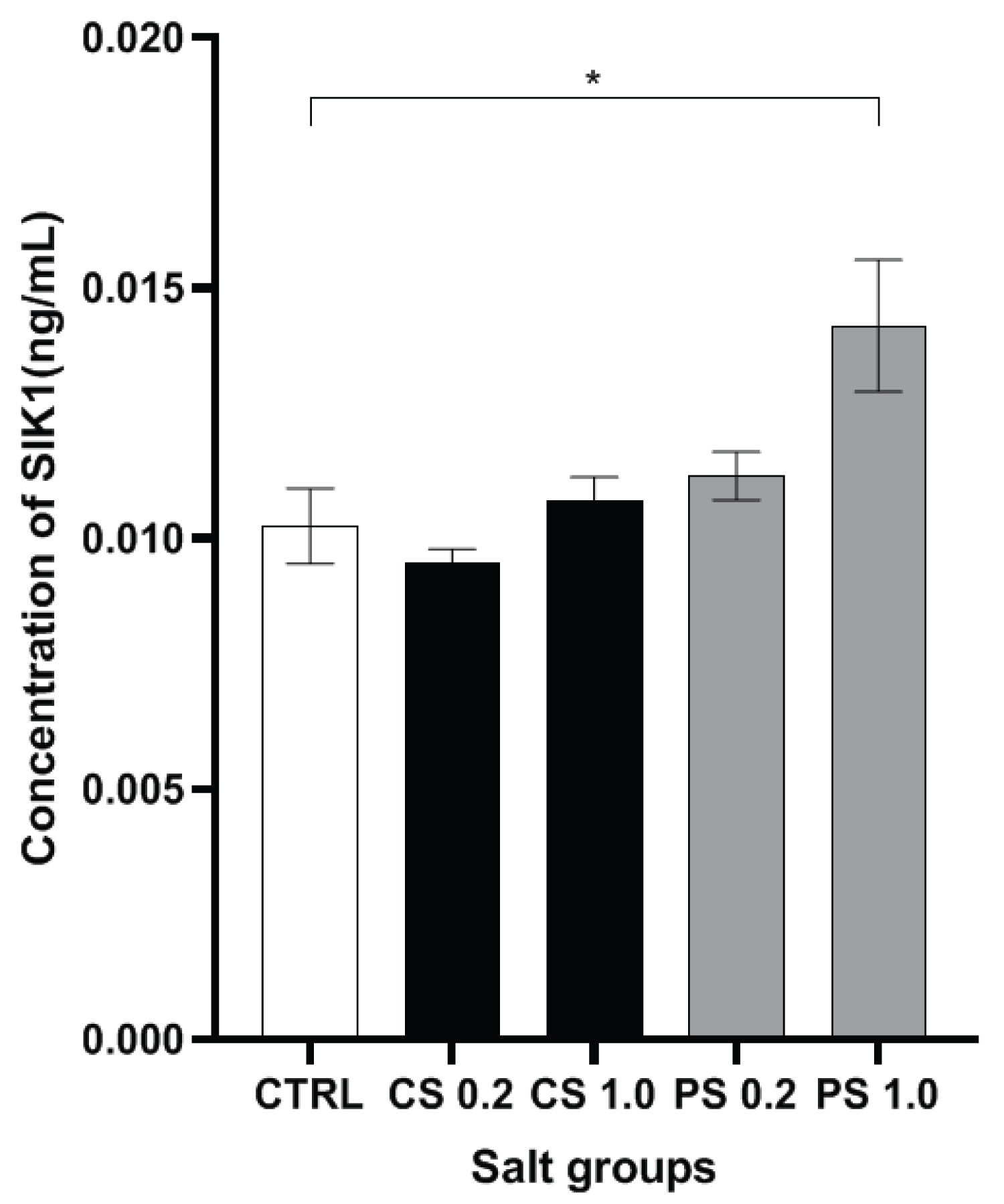
When the levels of SIK1 in the hippocampal tissue of the rats were assessed (Figure 3), a marginal increase in SIK1 levels was noted with 0.2 g/Kg treated Himalayan pink salt group. Importantly, a very significant elevation in the levels of SIK1 was noted with the 1.0 g/Kg pink salt group with reference to the control group. The greater SIK1 levels in the pink salt groups may be associated with a SIK1 mediated PP2A activation that regulates excessive tau phosphorylation and therefore prevents impairments in learning and memory as PP2A has been implicated in the pathogenesis of AD [14].
Measurement of phospho-Tau levels in the serum
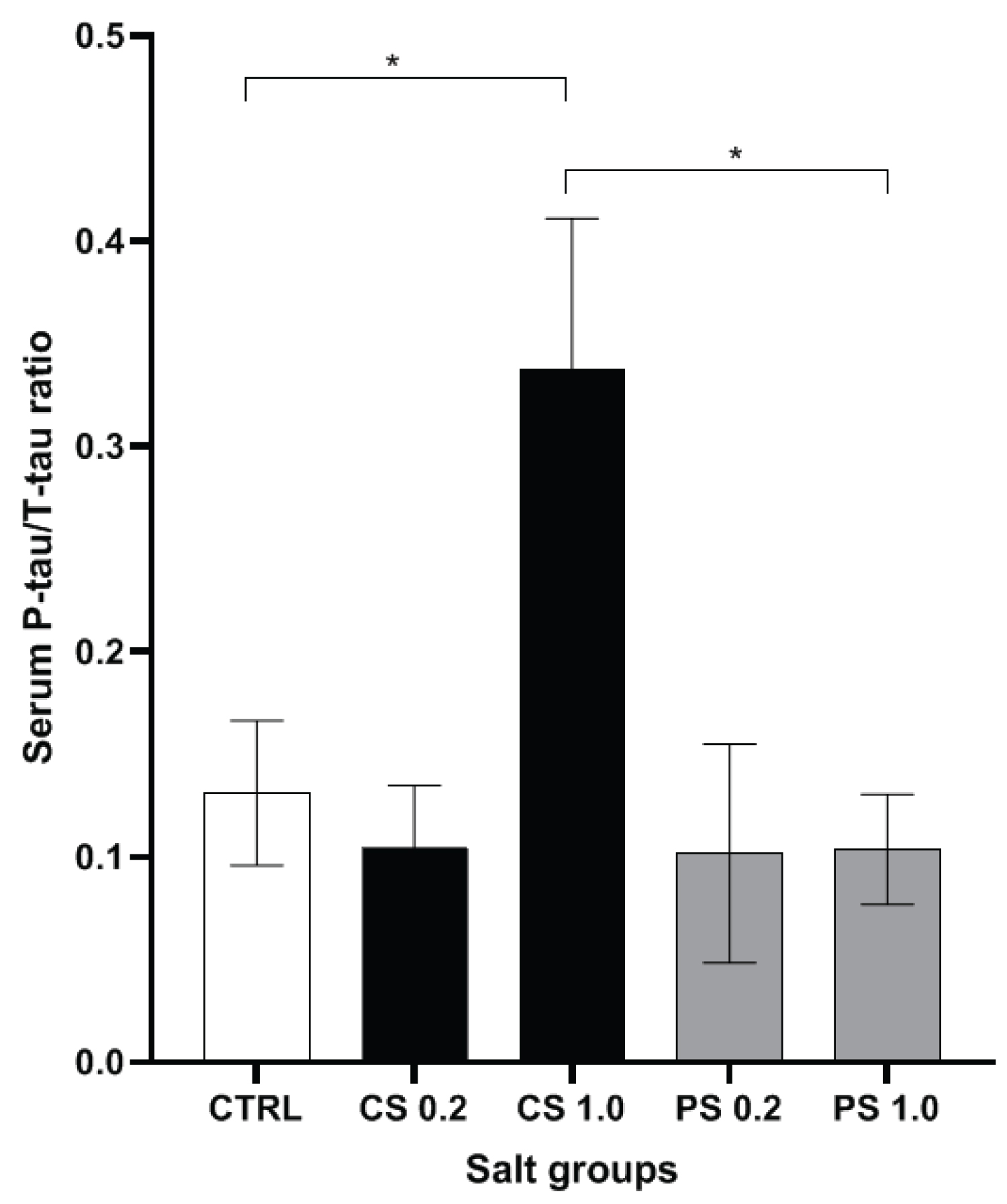
In the current study, the serum p-tau/total tau ratio was found to be significantly highest in the 1.0 g/Kg common salt treated group (Figure 4). Further, the results suggest the apparent lack of influence in promoting pathologically relevant hyper phosphorylation of tau by additional consumption of these salts for 12 weeks at 0.2 g/Kg body weight. It is very encouraging to note that, particularly, Himalayan pink salt did not elicit this undesirable increase in hyper phosphorylation even at 1.0 g/Kg body weight limit. In the previously mentioned assessment of NO metabolite levels in the serum, 1.0 g/Kg common salt was found to have the lowest nitrate/nitrite ratio indicating deficit in the NO production by e NOS. A recent study has reported that in mice fed on high salt diet, nitric oxide deficiency reduces nitrosylation of calpain. This results in its activation which in turn activates the enzyme cyclin-dependent kinase that leads to tau hyperphosphorylation [5]. Thus, the higher p-tau/tau ratio shown by the 1.0 g/Kg common salt group maybe explained to be as the result of tau phosphorylation occurring as a downstream result of endothelial dysfunction caused by excessive salt administration to the rats.
Discussion
Evidence from epidemiological studies support the hypothesis that cognitive decline is associated with modifiable lifestyle factors and its relation to diet is an area under intense research as this could open up new avenues for its prevention [15]. Existing results about the effect of dietary salt on cognitive function in humans are mixed. Assessment of the measures of pre-dementia or mild cognitive impairment in hypertensive women aged 65-79 or on antihypertensive medication showed a strong association of hypertension with cognitive impairment suggesting the involvement of salt-mediated vascular dysfunction as the underlying cause of cognitive impairment [16]. Cross-sectional studies conducted in a community sample comprising of 925 adults aged 50-96 reported that a lower dietary salt intake was associated with impaired executive functioning as assessed by Trails-B task and showed poorer performance on the Mini mental state examination (MMSE) [17]. Two prospective studies in aged adults conducted by assessment via food frequency questionnaires and MMSE reported no association between dietary salt intake and cognitive decline [18]. This calls for further research on the effects of dietary salt on cognitive decline under more tightly controlled conditions [4].
With respect to studies using animal models, mice fed for 7-28 days with high salt chow (> 4% NaCl) resulted in hypertension-independent blood brain barrier (BBB) disruption by downregulation of the tight junction protein ZO-1 and p38/MAPK-SGK1 dependent pathway [19]. A recent study examining the effect of dietary salt on cerebrovascular function and cognition showed that high-salt diet (4-8% NaCl which is equivalent to about 8-16 times the salt content of the normal mouse chow) for two to six months leads to cerebral endothelial dysfunction in the mice without causing hypertension and without affecting neurovascular coupling [20]. Further, a decreased NO production with decreased nitrosylation of calpain (regulator of Cdk5) and subsequent activation of Cdk5 was noted. This resulted in tau phosphorylation thus providing evidence that tau accumulation mediates memory impairments occurring as a result of high salt diet rather than cerebral hypoperfusion [5].
In the current study, the effect of chronic administration of 5-10 fold higher concentrations above the recommended daily intake of Himalayan pink salt with regard to its influence on spatial memory in comparison with that of common salt was evaluated using rat as an experimental model. The Himalayan rock salt, mined from the sedimentary rocks in the North-western Himalayan ranges, with its unique pinkish colour has gained popularity in households as well as haute cuisine. From examination of the chemical profile, this salt has been found to be richer in minerals in comparison to the common table salt [21]. In-vitro studies conducted with Himalayan rock salt have revealed that it has significant antioxidant potential, and this could be attributable to the presence of minerals such as iron, magnesium, and calcium [11].
High salt diet (HSD) induced endothelial dysfunction in both cerebral and systemic vasculature has long been known to be caused by a nitric oxide deficit [22]. Though most of the research so far has elucidated the causal mechanisms of salt-induced systemic hypertension, recent studies have focussed on the cerebro-endothelial dysfunction that occurs independent of changes in blood pressure. HSD has been found to produce an increase in circulating levels of the pro-inflammatory cytokine IL-17 due to the polarisation and differentiation of the regulatory T lymphocytes TH17 in the small intestine of HSD rodent models. Increased IL-17 levels in turn leads to decrease in NO production by an inhibitory phosphorylation of the endothelial nitric oxide synthase (e NOS) enzyme [20]. The results presented in this study (Figure 1) are in support of the available literature that high salt adversely affects endothelial homeostasis with implications for cerebral perfusion and cognitive function [5]. It is of interest to note that, chronic administration of very high concentration of upto 1.0 g/Kg body weight of Himalayan pink salt did not reduce the serum nitrate/nitrite ratio. Further, the higher nitrate/nitrite ratio noted in this experimental group in comparison to the control group (though not statistically significant) is indicative of an enhancing effect of Himalayan pink salt on NO production.
In Barnes maze task, significant reduction in latency and number of errors made by 0.2 g/Kg pink salt treated group (Figure 2) may be attributed to the reported antioxidant potential of Himalayan pink salt [11] thereby preventing hippocampal oxidative stress in the group. Further, the nitrate/nitrite ratio of the group as observed previously, was comparable to that of the control group indicating that the endothelial function was unaffected (normal NO production levels) despite the supra physiologic salt concentration that was administered. This has possible implications in prevention of possible learning impairment owing to a salt-induced cerebral hypoperfusion [5].
Salt-inducible kinases (SIK), are expressed as three isoforms, viz., SIK1, SIK2, and SIK3.Among them, SIK2 and SIK3 exhibit ubiquitous, constitutive expression in human tissues. In contrast, SIK1 mRNA expression is under the regulation of multiple stimuli such as high salt diet, adrenocorticotropin hormone, glucagon, circadian rhythms, and excitable cell depolarization [23]. SIK1 is also present in rat brain and upon depolarization, mRNA levels of SIK1 have been demonstrated to increase several fold in the hippocampus and cortex [24]. Increased SIK1 expression in rodent brain tissue upon exposure to alcohol has been shown to regulate alcohol-induced neuroinflammation [25]. Increase in intracellular sodium concentration has been shown to activate SIK1 as a part of the sodium-sensing network in Ca2+/Calmodulin kinase dependent manner in mammalian cells [26]. SIK1 is homologous to the SOS2 protein which is upregulated during salt stress in plants acting via calcium responsive kinase pathways [27]. SIK-1 is constitutively associated with the Na+/K+-ATPase complex. It is activated by increase in calcium via the Na+/Ca2+ exchanger upon elevation in intracellular sodium concentration. SIK-1 phosphorylates PME-1 of the PP2A/PME-1 complex and causes its dissociation from it. Thus, released active form of PP2A can dephosphorylate downstream substrates including the hyperphosphorylated AD-associated tau. The statistically significant increase in SIK1 levels in the pink salt treated experimental groups (Figure 3) suggest that SIK1 may be involved in the sodium buffering as part of SIK1-Na+/K+-ATPase complex in the brain. SIK1-Na+/K+-ATPase complex has an important functional role as part of the choroid plexus in maintaining sodium homeostasis in the brain. The above results imply that the Himalayan pink salt may have a protective role in maintaining the integrity of SIK1 network and sodium homeostasis in the rat brain. In addition, SIK-1 phosphorylates PME-1 of the PP2A/PME-1 complex and causes its dissociation from it. Thus, released active form of PP2A can dephosphorylate downstream substrates including the hyperphosphorylated AD-associated tau.
The elevations in the levels of phosphorylated tau and total tau in the cerebrospinal fluid are well established hallmarks of AD [28]. Due to invasiveness and time consumption of CSF sampling in humans, serum or plasma levels of tau have been considered to be alternative reliable AD biomarkers [29]. Further, backing the utility of serum p-tau and t-tau as biomarkers for AD, association between tau levels in CSF and plasma have been reported [30]. With the background of the aforementioned study reports and an extrapolation from the available literature that spinal cord injury (SCI) model rats exhibit significant correlation of tau protein levels between CSF and serum [31], in the present study, p-tau and total tau protein levels were measured in rat serum and the ratio of p-tau to total tau was considered (Figure 4). The serum p-tau/tau levels in the 1.0 g/Kg Himalayan salt group was comparable with that of the control group and the 0.2 g/Kg concentration salt groups. The elevated SIK1 levels in the 1.0 g/Kg Himalayan pink salt group in the previously mentioned result can be associated with the reduction in p-tau/t-tau ratio in the group with possible interaction of SIK1 with PP2A enzyme involved in dephosphorylation activity as in the mechanism elucidated in the previous results.
Limitations of the study include the small sample size, lack of total antioxidant status analyses in the hippocampal homogenates and lack of the experimental animals. Present study was limited to Barnes maze task as a measure of hippocampus-driven spatial memory function test. Additional behavioural studies such as active place avoidance task with larger sample size are needed to strengthen the current findings. Another caveat of the present study is that lipid peroxidation/glutathione levels/activity of antioxidant enzymes (superoxide dismutase, catalase, glutathione peroxidise, glutathione reductase) in hippocampal homogenates were not measured. It has only been extrapolated from available literature regarding the antioxidant properties of Himalayan pink salt. Further research about the role of salt inducible kinases and their role in brain with regard to inflammation, gene expression, and neuronal plasticity alterations with respect to the changes in various ion concentrations can bring new perspective to the understanding of the effects of dietary salts on the brain. Detailed studies about the role of SIK1 in sodium homeostasis in the brain are warranted.
In summary, from the results of the above-described assessments, we may infer that the Himalayan pink salt is a superior dietary salt substitute in comparison to the common salt as is evidenced by its notable enhancement of performance of the animals in the learning and memory assessment tasks as well as its effect in protecting against endothelial dysfunction and regulation of phospho-tau levels. These results can set the ground for further detailed investigations into the properties and effects of pink salt with respect to cerebrovascular alterations and their possible role in mechanisms of tau pathology as well as cognitive deficits.
Acknowledgement
Financial assistance to SS from SERB, New Delhi, India [Grant no. EMR/2017/000849] is gratefully acknowledged.
Conflict of Interest
The authors declare no conflict of interest.
References
- Johnson C, Praveen D, Pope A, et al. (2017) Mean population salt consumption in India: A systematic review. J Hypertens 35: 3-9.
- World Health Organization (2010) Global status report on noncommunicable diseases.
- Mente A, O'Donnell M, Rangarajan S, et al. (2018) Urinary sodium excretion, blood pressure, cardiovascular disease, and mortality: A community-level prospective epidemiological cohort study. Lancet 392: 496-506.
- Kendig MD, Morris MJ (2019) Reviewing the effects of dietary salt on cognition: Mechanisms and future directions. Asia Pac J Clin Nutr 28: 6-14.
- Faraco G, Hochrainer K, Segarra SG, et al. (2019) Dietary salt promotes cognitive impairment through tau phosphorylation. Nature 574: 686-690.
- Chui HC, Ramirez-Gomez L (2015) Clinical and imaging features of mixed Alzheimer and vascular pathologies. Alzheimers Res Ther 7: 21.
- Armstrong R (2019) Risk factors for Alzheimer's disease. Folia Neuropathol 57: 87-105.
- Trieu K, Neal B, Hawkes C, et al. (2015) Salt reduction initiatives around the world - a systematic review of progress towards the global target. PLoS One 10: e0130247.
- Carapeto C, BrumS, Rocha M (2018) Which table salt to choose? J Nutr Food Sci 8: 3.
- Fayet-Moore F, Wibisono C, Carr P, et al. (2020) An analysis of the mineral composition of pink salt available in Australia. Foods 9: 1490.
- Chander V, Tewari D, Negi V, et al. (2020) Structural characterization of Himalayan black rock salt by SEM, XRD and in-vitro antioxidant activity. Sci Total Environ 748: 141269.
- Joseph SK, Vishnuvardhan, Sushmitha C, et al. (2020) Phospho-Tau antibody delivery using virus-like nanoparticles as carriers enhances spatial memory functions in tauopathy model rats. Alzheimers Dis Dement 4: 99-104.
- Barnes CA (1979) Memory deficits associated with senescence: A neurophysiological and behavioral study in the rat. J Comp Physiol Psychol 93: 74-104.
- Alonso AD, Grundke-Iqbal I, Barra HS, et al. (1997) Abnormal phosphorylation of tau and the mechanism of Alzheimer neurofibrillary degeneration: Sequestration of microtubule-associated proteins 1 and 2 and the disassembly of microtubules by the abnormal tau. Proc Natl Acad Sci 94: 298-303.
- Caracciolo B, Xu W, Collins S, et al. (2014) Cognitive decline, dietary factors and gut-brain interactions. Mech Ageing Dev 136-137: 59-69.
- Haring B, Wu C, Coker LH, et al. (2016) Hypertension, dietary sodium, and cognitive decline: Results from the women's health initiative memory study. Am J Hypertens 29: 202-216.
- Rush TM, Kritz-Silverstein D, Laughlin GA, et al. (2017) Association between dietary sodium intake and cognitive function in older adults. J Nutr Health Aging 21: 276-283.
- Nowak KL, Fried L, Jovanovich A, et al. (2018) Dietary sodium/potassium intake does not affect cognitive function or brain imaging indices. Am J Nephrol 47: 57-65.
- Zhang T, Fang S, Wan C, et al. (2015) Excess salt exacerbates blood-brain barrier disruption via a p38/MAPK/SGK1-dependent pathway in permanent cerebral ischemia. Sci Rep 5: 16548.
- Faraco G, Brea D, Garcia-Bonilla L, et al. (2018) Dietary salt promotes neurovascular and cognitive dysfunction through a gut initiated TH17 response. Nature Neurosci 21: 240-249.
- Drake SL, Drake M (2011) Comparison of salty taste and time intensity of sea and land salts from around the world. Journal of Sensory Studies 26: 25-34.
- Boegehold MA (2013) The effect of high salt intake on endothelial function: reduced vascular nitric oxide in the absence of hypertension. J Vascular Res 50: 458-467.
- Wein MN, Foretz M, Fisher DE, et al. (2018) Salt-inducible kinases: Physiology, regulation by cAMP, and therapeutic potential. Trends Endocrinol Metab 29: 723-735.
- Feldeman JD, Vincian L, Crispino M, et al. (2000) The salt-inducible kinase, SIK, is induced by depolarization in brain.J Neurochem 74: 2227-2238.
- Zhang Y, Gao W, Yang K, et al. (2018) Salt-inducible kinase 1 (SIK1) is induced by alcohol and suppresses microglia inflammation via NF-?B signaling. Cell Physiol Biochem 47: 1411-1421.
- Sjöström M, Stenström K, Eneling, K, et al. (2007) SIK1 is part of a cell sodium-sensing network that regulates active sodium transport through a calcium-dependent process. ProcNatl Acad Sci 104: 16922-16927.
- Bertorello AM, Zhu JK (2009) SIK1/SOS2 networks: Decoding sodium signals via calcium-responsive protein kinase pathways. Eur J Physiol 458: 613-619.
- Nam E, Lee YB, Moon C, et al. (2020) Serum tau proteins as potential biomarkers for the assessment of alzheimer's disease progression. Int J Mol Sci 21: 5007.
- Chiu MJ, Chen YF, Chen TF, et al. (2014) Plasma tau as a window to the brain-negative associations with brain volume and memory function in mild cognitive impairment and early Alzheimer's disease. Hum Brain Mapp 35: 3132-3142.
- Tatebe H, Kasai T, Ohmichi T, et al. (2017) Quantification of plasma phosphorylated tau to use as a biomarker for brain Alzheimer pathology: Pilot case-control studies including patients with Alzheimer's disease and down syndrome. Mol Neurodegeneration 12: 63.
- Tang Y, Liu HL, Min LX, et al. (2019) Serum and cerebrospinal fluid tau protein level as biomarkers for evaluating acute spinal cord injury severity and motor function outcome. Neural Regen Res 14: 896-902.
Corresponding Author
Sarada Subramanian, Department of Neurochemistry, National Institute of Mental Health & Neurosciences, Bangalore, 560029, India, Tel: 91-80-2699-5165, Fax: 91-80-2656-4830, E-mail: sarada@nimhans.ac.in
Copyright
© 2021 Keerthana P, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.