Direct Conversion of Non-Functional Pluripotent Human Embryonic Stem Cells into Functional Somatic Elements Creates Scientific Paradigms to Address Key Challenges to Traditional Medicine and Biofabrication
Abstract
The limited capacity of neuron circuitry and myocardium for self-repair constitutes a significant challenge to traditional medicine for tissue and function restoration. Recent technology breakthroughs demonstrate that non-functional pluripotent human embryonic stem cells (hESC) can be directly and uniformly converted into a large supply of fate-restricted functional human neuronal or cardiomyocyte derivatives by small molecule induction, which not only constitute clinically representative progress in both human neuronal and cardiac therapeutic products for treating a wide range of incurable or hitherto untreatable neurological and heart diseases, but also offer manufacturing innovations for production scale-up and creation of replacement tissue and organ products. Such groundbreaking innovative technology platforms provide tissue and organ engineering paradigm for reconstitution or reconstruction of human central nervous system (CNS) or heart-related models or replacement tissue and organ products, which will pave the way for further development of cutting-edge automated high-content systems for systematic functional assembly of the in vitro replacement tissues and organs from pluripotent hESC in a 3D setting that reflect the characteristics and function of the in vivo human organ system, enabling automated high content and high-throughput analysis of the CNS or heart circuitries and dynamics, and systems developmental biology modeling of the complex human development and disorders. To date, the pluripotent hESC remain as the readily available source for genetically-stable human pluripotent stem cells (hPSC) with full-developmental potential in deriving somatic elements for human tissue and function restoration. Future efforts to relax policy and allocate funding to support such hESC research for the development and application of innovative technologies to address key challenges to traditional medicine and biofabrication will have a transformative impact on translational research priority, potentially shifting current research and clinical practices, and creating new scientific paradigms for CNS and heart repair.
Keywords
Regenerative medicine, Stem cell technology, Tissue-engineering, Cell therapy, Product manufacturing, Neurological disorder, Heart disease
Introduction
To date, the need to restore vital tissue and function for a wide range of neurological and heart diseases remain a daunting challenge to the conventional mode of drugs and treatments. The successful derivation of pluripotent human embryonic stem cell (hESC) lines from the in vitro fertilization (IVF) leftover embryos, which are otherwise destined for destruction, has brought one of the major breakthroughs of life sciences [1-5]. The hESC, derived from the inner cell mass (ICM) or epiblast of human blastocyst, are genetically stable with intrinsic capability for unlimited expansion and differentiation into all the somatic elements, proffering a inexhaustible nature source of human pluripotent stem cells (hPSC) for in vitro derivation of a large supply of disease-targeted human somatic cells that are restricted to the lineage in need of repair [1,2]. Therefore, they have been regarded as an ideal source to provide an unlimited supply of clinically-relevant functional human cells to heal the damaged or lost tissues that have naturally limited capacity for self-repair, such as the human brain and heart. Over the past decade, remarkable advancements on research of hPSC, including hESC and induced pluripotent stem (iPS) cells (Table 1), not only open up the future perspectives to resolve worldwide major health problems challenging to traditional medicine, such as brain disorders and heart diseases, but also offer powerful tools for systematic functional assembly of the in vitro replacement tissue and organ products at a commercial scale for regenerative or replacement therapies as well as for automated high content high-throughput analysis and modeling of the complex human development and disorders.
Technical Barriers of the Conventional Mode of Drugs and Treatments
Challenges to traditional medicine in addressing unmet medical needs
The neuron circuitry of the human brain and spinal cord of the central nervous system (CNS) has very limited capacity for self-repair, which constitutes a significant challenge to traditional medicine for nerve tissue and neurological function restoration in seeking cures for a wide range of neurological disorders. Given the limited capacity of the CNS for self-repair, transplantation of human neuronal stem cell therapy derivatives represents a promising therapeutic approach closest to provide a cure to restore the damaged or lost nerve tissue and function for neurological disorders. However, to date, the available sources for clinically-suitable engraftable human stem/progenitor cells with adequate neurogenic potential remain lacking, which has been not only the major setback in developing safe and effective cell-based neuronal therapies for regenerating the damaged or lost CNS structure and circuitry in a wide range of neurological disorders, but also the major bottleneck for CNS tissue engineering. The traditional sources of engraftable human stem cells with neural potential for transplantation therapies have been multipotent human neural stem cells (hNSC) isolated directly from the human fetal neuroectoderm or CNS. However, conventional cell therapies based on primary CNS tissue-derived hNSC have encountered supply restriction and difficulty to use in the clinical setting due to their declining plasticity with aging and limited expansion ability, making it difficult for production scale up in clinical translation, thus restricting tissue-resident primary hNSC as an adequate source for graft materials [5-7]. The hPSC have been regarded as an ideal source to derive an unlimited supply of large-scale well-characterized stem/progenitor/precursor cells in vitro for cell-based therapies and tissue engineering. Although hNSC have traditionally been isolated directly from the CNS (primary hNSC), the field is becoming more adept at deriving hNSC in vitro from hPSC (secondary hNSC), most classically exemplified by the hESC. The major challenge in using hPSC for cell-based therapy and tissue engineering is to produce a large and uniform population of lineage-committed transplantable cells in vitro. Although neural lineages appear at a relatively early stage in differentiation, only very small portion of hPSC (< 5%) undergo spontaneous differentiation into neurons under cultivation [2,5-7]. The neural grafts derived secondarily from hPSC in vitro through conventional multi-lineage differentiation yielded neurons at a low prevalence following engraftment [5-11]. Such low yields were not only insufficient for regeneration or reconstruction of the damaged CNS, but also accompanied by unacceptably high incidents of teratoma and/or neoplasm formation, raising considerable safety concerns [5-11]. Similar to CNS-derived primary hNSC, hPSC-derived secondary hNSC are neuroepithelial-like nestin-positive cells that spontaneously differentiate into a heterogeneous population of mixed cell types containing undifferentiated cells, different subtypes of neurons (5-15%), astrocytes, and oligodendrocytes in vitro and in vivo [5-11]. Despite some beneficial outcomes, traditional sources of hNSC, either isolated primarily from CNS tissues in vivo or derived secondarily from hPSC in vitro, appeared to exert their therapeutic effect primarily by their non-neuronal progenies through producing trophic and/or neuro-protective molecules to rescue endogenous host neurons or through enhancing endogenous repair, but not related to graft-dependent regeneration or repair [5-11].
Similarly, the functional contractile muscle cells of the heart-known as cardiomyocytes-have very limited capacity for self-repair after birth, which necessitates cell-based therapy for myocardium regeneration and contractile function restoration in seeking treatments or cures for heart disease and failure. However, to date, the available sources for clinically-suitable engraftable human stem/progenitor/precursor cells or cardiomyocytes with adequate myocardium regenerative potential remain lacking, which has been not only the major setback in developing safe and effective cell-based heart therapies for myocardium tissue and contractile function restoration in cardiovascular disease, but also the major bottleneck for myocardium tissue engineering [5,7-11]. In the adult hearts, cardiomyocytes are terminally differentiated and unable to regenerate. Despite numerous reports about small quantities of cell populations expressing stem/progenitor cell markers identified in the adult hearts, growing evidences indicate that they are not genuine heart cells and give rise predominantly to non-functional smooth muscle cells rather than functional contractile cardiomyocytes [7-11]. There is no evidence that stem/precursor/progenitor cells derived from other tissues or sources, such as bone marrow, cord blood, umbilical cord, mesenchymal stem cells, placenta, or fat tissue, are able to give rise to the contractile heart muscle cells following transplantation into the heart [7-11]. During the past decade, the hPSC have become a vital human cardiac cell source for myocardial tissue engineering and myocardium repair. However, in hPSC-differentiating multi-lineage embryoid bodies, only a very small fraction of cells (< 4%) spontaneously differentiate into cardiomyocytes [7-11]. Even though such hPSC-derived immature cardiomyocytes can be enriched to attenuate the progression of heart failure in an acute myocardial infarction model up to 12 weeks, the grafts generated by cell transplantation have been small and insufficient to restore heart function or to alter adverse remodeling of chronic infarcted models following transplantation [7-11]. Functional enhancement in preclinical animal models by such hPSC-derived cardiomyocytes has been limited to mid-term at most, equivalent to perhaps a few months in humans, and there is no evidence that the underlying mechanism depends on the contractile properties of the transplanted human cells [7-11].
A host of major health problems result in tissue or organ damage or loss, thus functional impairment or loss, which cost worldwide healthcare systems $ trillions every year [7-12]. Despite they are leading causes of death or permanent disability, yet, there is no effective treatment available for those devastating and life-threatening diseases [7-12]. The limited capacity of those vital human tissues, such as neuron circuitry of the brain and myocardium of the heart, for self-repair mandates the designated cell therapy product to have adequate efficacy or therapeutic/pharmacologic activity to regenerate the functional and structural tissue element that has been damaged or lost, such as neuron circuitry of the brain and contractile myocardium of the heart [7-12]. Therefore, the chaperone utility of traditional stem cells to produce trophic or protective molecules to rescue endogenous dying host neurons or cardiomyocytes that can simply be achieved by injection of a compound drug of molecular entity, such as trophic or growth factors, is insufficient to designate them as a cell therapy product or stem cell medicine [7-17]. Nevertheless, the high toxicities of trophic or growth factors in clinical trials have been well-documented [7,8]. Novel solutions are crucial for developing safe and effective cell-based therapies for regeneration of the damaged or lost CNS or myocardium structure and circuitry evidenced in various neurological and heart diseases.
Lack of scalable replacement human cell source for neurons and cardiomyocytes
In the neuroscience and heart fields, the lack of scalable clinically-suitable human cell source adequate for neuron circuitry and myocardium regeneration is a major drawback not only for achieving damaged or lost human nerve and myocardium tissue regenerative solutions in the clinical setting, but also for CNS and myocardium tissue engineering [7-17]. In addition, a substantial number of neurological disorders, including stroke, traumatic brain injury (TBI), spinal cord injury (SCI), result in extensive CNS nerve tissue damages or losses that require the development and application of novel CNS tissue engineering or biofabrication approaches in order to achieve effective regeneration or repair of the damaged or lost human neuronal tissues and circuitries, thus lead to a dramatic neurological function improvement or recovery in the clinical setting. In cardiac cell therapy, current cell delivery methods to the damaged heart, by injection of cells either directly into the infarcted region or through the coronary circulation, are inefficient [13-17]. In addition, arrhythmogenesis is a potential risk in cell-based heart therapy [13-17]. For end-stage heart failure, stem/progenitor-cell-mediated cellular regenerative approach cannot be used as an alternative approach to heart transplantation [13-17]. Those shortcomings provoke the need to develop tools for 3D heart assembly from hESC cardiomyocyte derivatives as the replacement heart in order to provide alternative treatment options to address acute shortage of donor organs in heart transplantation [7,8,13-17]. In case of successful heart transplantation from suitable donor organ, it requires life-long immune suppression that is often associated with serious side effects. The hESC and their derivatives are considerably less immunogenic than adult tissues [7-17]. It is also possible to bank large numbers of human leukocyte antigen isotyped hESC lines so as to improve the likelihood of a close match in clinical situations [7-17]. Therefore, the frontiers of medicine of making CNS- and heart-related tissue and organ products from hESC neuronal and cardiomyocyte derivatives, respectively, will not only provide 3D human CNS and heart models for systems developmental biology studies of the human CNS and heart circuitries and dynamics, but will also translate to clinical studies for organ-replacement therapies in order to meet some of the critical medical challenges resulted from acute shortage of donor organs and immune rejection in transplantation, potentially shifting current clinical practice paradigms for CNS and heart repair.
The valley of death
Traditional drug development usually starts with drug leads discovered in non-human simple model organisms, thus requires lengthy and costly both demonstration in animal model testing and establishment of proof-of-concept and safety in human trials. Many of the biological pathways and mechanisms of non-human lower-organism or animal model systems do not reflect the complexity of humans, thus have little implications for the prevention and treatment of human diseases in the clinical setting. As a result, millions of drug leads have vanished before even reach clinical trials, and for few lucky ones, have encountered the very high drug failure rate in human trials. Among those very few drugs that eventually obtained their market approvals, there were not any cures, or even meaningfully effective treatments, for Alzheimer's disease, Parkinson's disease, stroke, spinal cord and brain injuries, heart disease and failure, or a host of other disorders that destroy lives. The difficulty of crossing the gulf between finding a promising new agent and demonstrating its safety and efficacy in humans in biomedical research has been referred as "the valley of death". It is the pounding consequence of a vast amount of Federal and private investments only go to maintain the status quo of mainstream biomedical research in the non-human model organisms or systems. Without a readily accessible and effective human model system to unlock the mysteries of human development and disorders, the road of desperately seeking cures has become all but a dead end to real world remedy.
Shortcomings of conventional hpsc multi-lineage differentiation approaches
The intrinsic ability of a hESC of both unlimited self-renewal and differentiation into neural or cardiac elements makes it a practically inexhaustible source of replacement cells for CNS or myocardium circuitry and function restoration. As neurological and heart diseases incur exorbitant costs on the healthcare system worldwide, there is a strong focus on translating hESC research to provide newer, more efficient solutions for these unmet therapeutic needs. However, the hPSC themselves are unspecialized non-functional cells that cannot be used for therapeutic applications. It has been recognized that hPSC must be turned into fate-restricted specialized functional cells, a process known as differentiation, before use for cell therapy. A major bottleneck for clinical translation of hESC biology is to enable a well-controlled and efficient induction of hESC exclusively and uniformly to a specific clinically relevant lineage. Conventional hESC differentiation approaches rely on germ-layer induction of pluripotent cells, resulting in uncontrollable and unpredictable simultaneous multi-lineage differentiation that yields embryoid bodies or aggregates [5-11]. Embryoid bodies or aggregates consist of a mixed population of cell types of three embryonic germ layers, among which only a very small fraction of cells display targeted differentiation [5-11]. Those hESC multi-lineage differentiation approaches are laborious, costly, and need time-consuming purification, isolation, or selection procedures to generate only a small quantity of desired cells, impractical for commercial and therapeutic applications [5-11]. The hESC-derived cellular products through conventional multi-lineage differentiation approaches consist of a heterogeneous population of mixed cell types, including fully differentiated cells, high levels of various degrees of partially differentiated or uncommitted cells, and low levels of undifferentiated pluripotent cells, often resulting in poor performance and high tumor risk of such cell derivatives and tissue-engineering constructs following transplantation [5-11]. How to channel the wide differentiation potential of hPSC efficiently and predictably to a neural or cardiac phenotype has been a major challenge for both developmental study and clinical translation in the neuroscience and heart fields. In addition, undefined foreign or animal biological supplements and/or feeder cells that have typically been used for the isolation, expansion, and differentiation of hPSC make such cell derivatives unsuitable for clinical applications.
Emerging Innovative Technologies of Stem Cell Medicine
PluriXcel human stem cell technology platforms
In regenerative medicine, the clinical translation of stem cell research and innovation capabilities demonstrated by hESC investigations holds huge promise for treating major human diseases challenging traditional medicine. Due to the prevalence of the CNS and heart diseases worldwide, there is a large unmet healthcare need to develop hESC-based therapeutic solutions to provide optimal regeneration and reconstruction treatment options for tissue and function restoration. Millions of people are pinning their hopes on hESC research to provide novel and effective therapeutic solutions to overcome both the theoretical and technical barriers of conventional mode of treatments or drugs for such major health problems.
Recent progresses on emerging technologies of regenerative medicine provide groundbreaking innovative technology platforms the PluriXcel human stem cell technology platforms-enabling large-scale production of high quality clinical-grade hESC lines and direct conversion of such nonfunctional pluripotent cells by small molecule induction into large commercial scales of high quality clinical-grade functional human neuronal or cardiomyocyte derivatives [5-27]. Such progresses not only constitute clinically representative breakthroughs in both human neuronal and cardiac therapeutic products for treating a wide range of incurable or hitherto untreatable neurological and heart diseases, but also offer manufacturing innovations for production scale-up and creation of replacement tissue and organ products [7,8]. This human stem cell technology is based on the discovery that pluripotent hESC maintained under defined culture conditions can be directly and uniformly converted into functional human neuronal or cardiomyocyte cell therapy derivatives by small molecule induction. Direct conversion of nonfunctional pluripotent cells into clinically-relevant lineages enables large scale manufacturing of clinically-suitable functional human neuronal or cardiac therapeutic products from hPSC as stem cell medicine to provide the next generation of cell-based therapeutic solutions for a wide range of neurological and cardiovascular diseases, thus addresses crucial technical barriers necessary before bringing stem cell therapy to the clinic for unmet medical needs.
PluriXcel technology platforms provide proprietary clinical-grade translating capabilities to address key challenges to traditional medicine and biofabrication, currently offering (1) PluriXcel-DCS Platform for creation of clinical-grade human stem cell lines/products/banks, which provides defined culture systems (DCS) for well-controlled efficient derivation, maintenance, and differentiation of high quality clinical-grade stable hPSC lines, and for creating current good manufacturing practice (cGMP)-compatible/quality clinical-grade master/working cell banks (MCB/WCB) and tool boxes; (2) PluriXcel-SMI-Heart Platform for scale-up production of high quality contractile human cardiomyocytes at a commercial quantity necessary for myocardial tissue biofabrication and heart regenerative therapies, which provides scale-up cGMP manufacturing capability for high efficient direct conversion of non-functional hPSC into a large commercial scale of high quality clinical-grade human heart cell therapy derivatives adequate for development of safe and effective stem cell therapies for myocardium repair, and for creation of replacement heart tissue/organ products or models targeting heart disease and failure; (3) PluriXcel-SMI-Neuron Platform for scale-up production of high quality human neurons at a commercial quantity necessary for CNS tissue biofabrication and neuronal regenerative therapies, which provides scale-up cGMP manufacturing capability for high efficient direct conversion of non-functional hPSC into a large commercial scale of high quality clinical-grade human neuronal cell therapy derivatives adequate for development of safe and effective stem cell therapies for a wide range of neurological disorders, and for creation of CNS-related replacement tissue/organ products or models.
Plurixcel technology breakthroughs
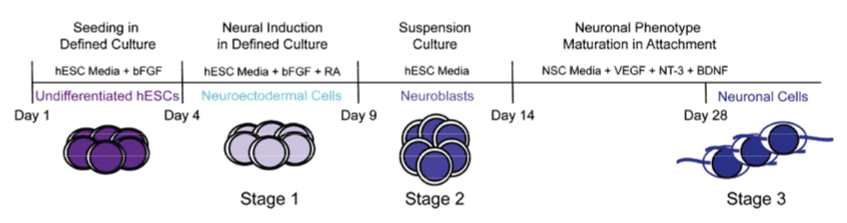
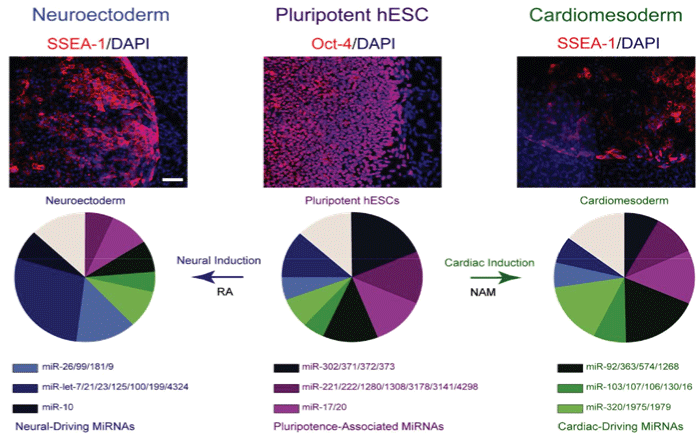
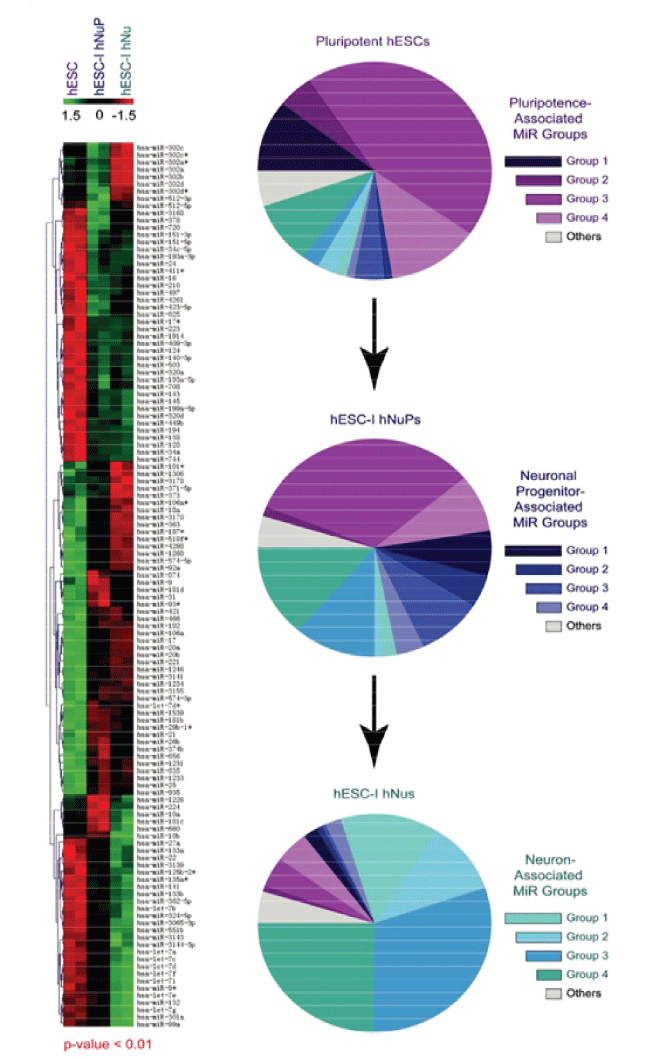
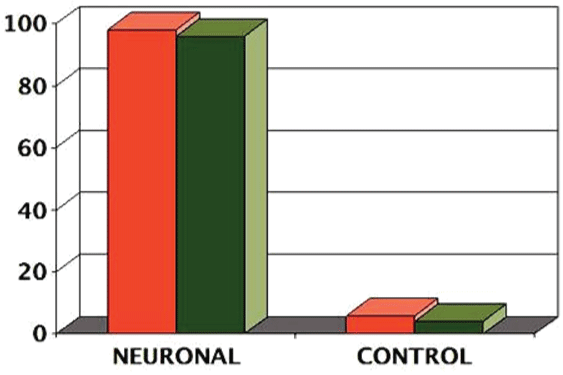
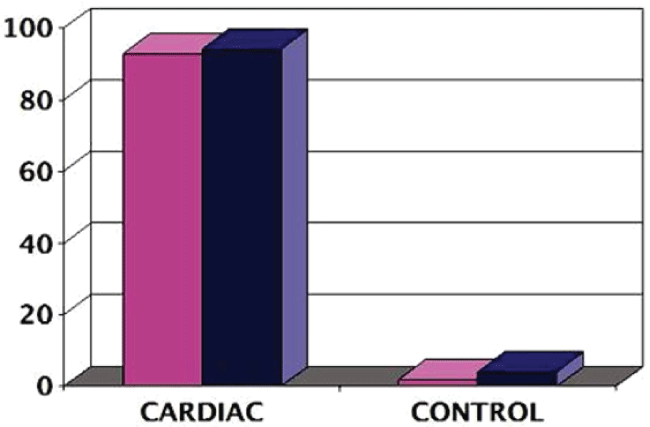
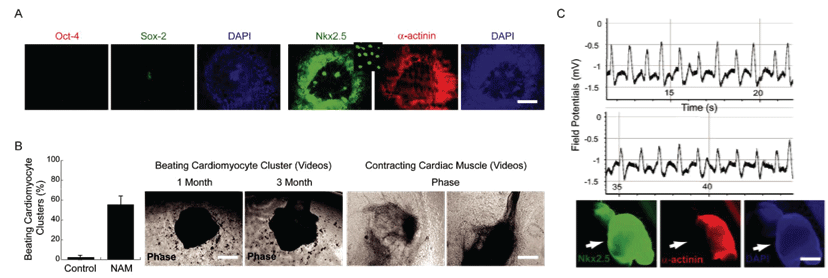
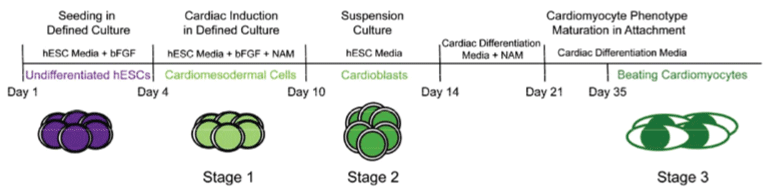
PluriXcel technology breakthroughs render well-controlled high efficient neuronal or cardiac lineage-specific conversion directly from the pluripotent state of hESC by small molecule induction, which open the door for human neural and cardiac tissue or organ engineering and regeneration as well as to investigate molecular neurogenesis and cardiogenesis in human embryonic development using powerful in vitro model systems [7,8]. Such breakthrough is one of the major milestones towards human trials of hESC therapy derivatives, demonstrating nonfunctional pluripotent hESC can be directly and uniformly converted into a specific clinically-relevant functional lineage by small molecule induction [5-27]. In particular, retinoic acid (RA) was identified as sufficient to induce the uniform specification of neuroectoderm direct from the pluripotent state of hESC maintained under the defined culture, without going through a multi-lineage embryoid body stage, and trigger a cascade of neuronal lineage-specific progression to human neuronal progenitors (Xcel-hNuP or hESC-I hNuP) and neurons (Xcel-hNu or hESC-I hNu) of the developing brain in high efficiency, purity, and neuronal lineage specificity by promoting nuclear translocation of the neuronal specific transcription factor Nurr-1 [5-12,20-25] (Figure 1, Figure 2, Figure 3, Figure 4 and Figure 5). Similarly, formulation of minimal essential defined conditions for hESC rendered small molecule nicotinamide (NAM) sufficient to induce the uniform specification of cardiomesoderm direct from the pluripotent state of hESC by promoting the expression of the earliest cardiac-specific transcription factor Csx/Nkx2.5 and triggering progression to cardiac precursors (Xcel-hCardP or hESC-I hCardP) and beating cardiomyocytes (Xcel-hCM or hESC-I hCM) in high efficiency [7-17,20,25-27] (Figure 2, Figure 6, Figure 7 and Figure 8). Nuclear translocation of NAD-dependent histone deacetylase SIRT1 and global chromatin silencing lead to small molecule-directed hESC cardiac fate determination or cardiomesoderm specification, while silencing of pluripotence-associated microRNA (miRNA) hsa-miR-302 family and drastic up-regulation of neuroectodermal Hox miRNA hsa-miR-10 family lead to small molecule-directed hESC neural fate determination or neuroectoderm specification [7-12,20,22-25] (Figure 2 and Figure 3). Small molecule induction transforms non-functional pluripotent hESC directly and uniformly into neuronal lineage-committed embryonic progenitor cells (Xcel-hNuP) [5-12,20-25]. The Xcel-hNuP yielded exclusively neurons (Xcel-hNu) in a drastically increased efficiency (> 95%) in vitro and in high abundance following transplantation into the brain, distinctly different from the prototypical neuroepithelial-like hNSC either isolated from the CNS tissues in vivo or derived from hPSC in vitro [5-12,20-25] (Figure 4 and Figure 5). Similarly, small molecule induction transforms nonfunctional pluripotent hESC directly and uniformly into cardiomyocyte lineage-committed embryonic precursors (Xcel-hCardP) [7-17,20,25-27]. The Xcel-hCardP yielded contractile cardiomyocytes (Xcel-hCM) in high efficiency (> 90% expressing cardiomyocyte markers, > 50% beating), distinctly different from the prototypical epithelial-like human cardiac stem cells either isolated from tissues in vivo or derived from hPSC in vitro [7-17,20,25-27] (Figure 6 and Figure 7). Accessory cells (e.g., glial cells or smooth muscle cells) and inappropriate cells (e.g., undifferentiated cells, cytotoxic cells, and non-neural or non-cardiac cells) are undetectable by high-sensitive molecular and genome-wide analysis (e.g., miRNA profiling) [5-17,20-27] (Figure 3). Such technology breakthroughs demonstrate that non-functional pluripotent cells can be directly and uniformly converted into a large supply of fate-restricted functional human cell therapy derivatives [7-12]. Direct conversion of non-functional pluripotent cells into functional somatic cells dramatically increases the clinical efficacy of graft-dependent repair and safety of hPSC-derived cellular products, marking a turning point in stem cell medicine from current studies in animals towards human trials or first-in-human studies [7-12]. Medical and manufacturing innovations of PluriXcel technology platforms provide life scientists and clinicians with novel, efficient, and powerful resources and tools to address major health concerns, which will shape the future of healthcare by providing safe and effective stem cell medicine for human tissue and function restoration, and bringing new regenerative therapeutics into the market.
PluriXcel technologies overcome major bottlenecks for cns/heart repair and biofabrication
PluriXcel technology breakthroughs have demonstrated the direct therapeutic or pharmacologic capability of hESC therapy derivatives for human CNS and myocardium regeneration, thus have presented the hESC therapy derivatives as a powerful pharmacologic agent of cellular entity for a wide range of incurable or hitherto untreatable neurological and heart disorders. Visualizing the medicinal utility of hESC will have a transformative impact on translational research priority by presenting hESC as a viable and robust therapeutic strategy for a wide range of incurable or hitherto untreatable neurological and heart diseases challenging traditional medicine, potentially shifting current research and clinical practices, and creating new scientific paradigms for CNS and heart repair [7-12].
PluriXcel technology platforms pioneer next-generation manufacturing techniques for repairing and replacing cell, tissue, and organ products, providing tissue and organ engineering paradigm for further reconstitution or reconstruction of complex multi-cellular 3D human CNS- or heart-related models or replacement tissue and organ products. Such 3D hESC constructs can be used for rapid and high fidelity safety and efficacy evaluation of human therapeutic candidates or cell therapy products, thus lead to advances in technologies used in the regulatory review; and will be readily adaptable in CNS-or heart-related drug efficacy and toxicity testing; and for commercialization and therapeutic development of replacement CNS- or heart-related tissue and organ products. It will pave the way for further development of cutting-edge automated high-content systems for systematic functional assembly of the in vitro replacement tissues and organs from pluripotent hESC in a 3D setting that reflect the characteristics and function of the in vivo human organ system, enabling automated high content and high-throughput analysis of the CNS or heart circuitries and dynamics, and systems developmental biology modeling of the complex human development and disorders. As a result, the breakthroughs of PluriXcel technology have led to the historical launch of Presidential Initiatives, including the Brain Research through Advancing Innovative Neurotechnologies (BRAIN) Initiative and, recently, the Advanced Tissue Biofabrication Manufacturing Innovation Institute (ATB-MII) Initiative. Progress of such initiatives will allow us to face the Grand Challenge in biomedical research focused on unlocking the mysteries of humans in development and disorders that are critical to its ultimate goal of improving human health.
Unique Clinical-Grade Translating Capabilities of hESC in Addressing Key Challenges to Traditional Medicine
Newer and more efficient solutions uniquely address key industry challenges to traditional medicine
PluriXcel technology breakthroughs have addressed key industry challenges to traditional medicine, offering a stem cell drug of cellular entity capable of novel regenerative medicine activity associated with human nerve and myocardium tissue and function restoration [7-11]. Such newer and more efficient solutions for the therapeutic needs of a wide range of neurological and heart diseases brought by the regenerative functionality intrinsic to the pluri-potential of hESC cannot be replaced by any other stem cells, including artificially-reprogrammed cells or iPS cells [7-12] (Table 1). The safety of a cell therapy product is evaluated by whether it can retain a normal stable phenotype and karyotype for a long period of time and whether there is no tumor, inappropriate cell type formation, or abnormal regeneration following transplantation. Embryo-originated pluripotent hESC, which display high levels of expression of Oct-4, SSEA-4, Tra-1-60, and Tra-1-81, can maintain prolonged normal stable growth or self-renewal in non-hostile growth environments containing the essential developmental components that sustain hESC pluripotency and self-renewal [5-11,18,19]. However, so far, there is no evidence that pluripotent cells derived from other sources harboring adult nuclei by transcription-factor- or small-molecule-directed reprogramming or somatic cell nuclear transfer, such as iPS cells or pluripotent cells derived from cloned embryos, can maintain prolonged normal stable growth or self-renewal [5-11, 18,19].
The broad potential of pluripotent hESC is defined by an epigenome constituted of open conformation of chromatin [7-11,18,19,22-25]. The epigenome of embryo-originated hESC is not only pluripotent, but also incredibly stable and positive, as evident by that only the positive active chromatin remodeling factors, but not the negative repressive chromatin remodeling factors, can be found in the open epigenome of pluripotent hESC [7-11,18,19,22-25]. The openness of pluripotent epigenome differentiates the active pluripotency of normal stable hESC from the repressive pluripotency of abnormal cells, such as the iPS cells reprogrammed from adult cells, pluripotent cells derived from cloned embryos, and pluripotent embryonic carcinoma cells [7-11,19] (Table 1). Although pluripotent, the artificially-reprogrammed cells or iPS cells are made from adult cells, therefore, they carry many abnormal negative repressive chromatin remodeling factors and unerasable genetic imprints of adult cells that normal stable pluripotent hESC do not have [7-11,19] (Table 1). Somatic nuclear transfer and factor- or small-molecule-directed reprogramming are incapable of restoring the correct epigenetic pattern of pluripotent hESC, which accounts for abnormal gene expression, accelerated senescence, and immune-rejection following transplantation of reprogrammed cells [7-11,19] (Table 1). Current artificially reprogramming techniques have not been able to resolve the genomic abnormality and instability of reprogrammed, induced, transdifferentiated, dedifferentiated, or cloned adult cells, which are associated with high risk of cancerogenesis, thus have severely limited their utility as viable therapeutic approaches [7-11,19]. In fact, serious spontaneous mutations were recently identified in human trials of iPS cells, further raising constant safety concerns for iPS cells that are made with the same mechanisms of oncogenesis by introducing or activating oncogenes in adult cells, thus intrinsically prone to mutations and genomic instability that could largely compromise their translational value as a viable therapeutic approach [7,19].
Although pluripotent, human iPS cells reprogrammed from adult cells or fibroblasts may reside in different developmental stages, and may not respond to developmental or inducing signals the same way as the hESC do [7,8]. In fact, the same small molecules do not induce cardiomesodermal or neuroectodermal differentiation of human iPS cells, nor trigger their progression to cardiomyocytes or neurons, even though previous reports show small molecule inhibitors of Wnt signaling can promote human iPS cells and certain reporter engineered-hESC lines differentiate into cardiomyocytes [7,8] (Table 1). So far, although a vast sum of government and private investments have been spent on looking for adult alternates, such as reprogramming and transdifferentiation of fibroblasts or mature tissues, only human stem/precursor/progenitor cells derived from embryo-originated pluripotent hESC have demonstrated such cellular pharmacologic utility and capacity adequate for CNS and myocardium regeneration in pharmaceutical development of stem cell therapy for neurological and heart disorders [7-12]. Thus, the pluripotent hESC remain as currently the only available source for genetically-stable hPSC with full-developmental potential in deriving somatic elements for human tissue and function restoration [7-12].
PluriXcel technology is one of the major achievements for clinical translation of hESC research, offering currently the only available human cell sources in large quantities and high quality with adequate cellular pharmacologic utility and capacity to regenerate the neuron circuitries and contractile heart muscles, vital for the CNS and myocardium repair for a wide range of neurological and heart disorders in the clinical setting [7-12]. Traditional hPSC multi-lineage differentiation approaches are unpredictable, uncontrollable, unrepeatable, low-efficiency, phenotypic heterogeneity and instability, high risk of tumor and/or inappropriate cell type formation following transplantation, and require laborious, costly, and time-consuming purification or isolation procedures to generate only a small quantity of desired cells. PluriXcel approach overcomes the shortcomings of traditional hPSC multi-lineage differentiation approaches that are impractical for production scale-up necessary in commercial and clinical applications [5-11]. PluriXcel approach offers the benefits in efficiency, stability, safety, efficacy, and large-scale production of high quality clinical-grade human neuronal or cardiac cell therapy products in current cGMP facility for commercial and therapeutic uses over all other existing approaches [5-17,20-27].
Novel therapeutic approach fundamentally different from traditional strategies
PluriXcel technology provides a novel therapeutic approach, fundamentally different from traditional strategies, enabling the efficient production of a large commercial scale of clinical-grade high quality functional human neuronal or cardiomyocyte therapeutic derivatives adequate for neuron circuitry and myocardium regeneration and biofabrication by utilization of biologics-free defined culturing and small molecule induction at the pluripotent stage of hESC [7-17,20-27]. The hESC neuronal cell therapy derivatives of PluriXcel approach are homogenous populations of neuronal lineage-committed cells with a high degree of potency for regeneration or reconstruction of CNS neuron circuitry of the human brain and spinal cord, and contain no residual pluripotent cells and other cellular impurity of safety concerns, dramatically increasing the clinical efficacy of graft-dependent neuronal repair and safety of hESC-derived cellular products for scale-up regeneration in the clinical setting, distinctly different from traditional sources of hNSC either isolated from CNS tissues in vivo or derived from hPSC in vitro [5-12,20-25] (Figure 1, Figure 2, Figure 3, Figure 4 and Figure 5). The hESC cardiomyocyte derivatives of PluriXcel approach are homogenous populations of cardiomyocyte lineage-committed cells with a high degree of potency for myocardium regeneration or reconstruction, and contain no residual pluripotent cells or other cellular impurities of safety concerns, dramatically increasing the clinical efficacy of graft-dependent myocardial repair and safety of hESC-derived cellular products for scale-up regeneration in the clinical setting, distinctly different from traditional sources of human cardiac stem cells either isolated from the heart in vivo or derived from hPSC in vitro [7-17,20,25-27] (Figure 2, Figure 6, Figure 7 and Figure 8). PluriXcel technology platforms introduce stem cell medicine development and manufacturing innovations to achieve clinically-useful and therapeutically-viable levels of safety and efficacy in cell therapy product development, providing unique clinical-grade translating capabilities for moving stem cell research from current studies in animals into human trials or first-in-human studies.
PluriXcel technology is an unconventional and exceptionally innovative approach in neuronal or cardiomyocyte cell therapy product development and production scale-up. The novelties of PluriXcel approach include: (1) it enables well-controlled high efficient neuronal or cardiac lineage-specific differentiation direct from the pluripotent state of hESC by small molecule induction, without going through a multi-lineage embryoid body stage, fundamentally different from traditional hESC multi-lineage differentiation approaches using germ-layer induction of pluripotent cells; (2) it enables high efficient direct conversion of non-functional pluripotent cells into a large supply of functional neuronal or cardiac fate-restricted derivatives for repair or regeneration of neuron circuitry or myocardium in the clinical setting, a major milestone towards human trials or first-in-human studies of stem cell medicine; (3) it provides a adequate source of human neuronal or cardiomyocyte cell therapy products in high purity and large quantity for neuron circuitry or myocardium regeneration or repair in treating a wide range of neurological and heart diseases; (4) it provides a homogenous population of neuronal or cardiomyocyte lineage-committed cells, which contain no residual pluripotent cells and other cellular impurities of safety concerns, with a high degree of potency for human neuron circuitry or myocardium regeneration in vitro and in vivo, distinctly different from traditional sources of human stem cells either isolated from the tissues in vivo or derived from hPSC in vitro; (5) it dramatically increases the clinical efficacy of graft-dependent neuronal or myocardium repair and safety of stem cell therapy products, distinctly different from traditional sources of human stem cells that exert their therapeutic effect primarily by enhancing endogenous repair, but not related to regeneration or repair from the graft; (6) it provides scalable platforms to ensure a high degree of efficacy and safety of stem cell therapy products, thus robust clinical benefits leading to therapies, for treating major human diseases challenging traditional medicine, which represent the next generation of human cell therapy products, offering purity, large-scale production, high quality, safety, efficacy, and effectiveness for commercial and therapeutic uses over all other existing human cell sources; (7) it provides scale-up cGMP manufacturing capabilities or techniques for production of large quantities of high quality clinical-grade stem cell therapy products to support clinical trials and tissue or organ engineering, improving the availability, reproducibility, accessibility, and standardization of manufacturing materials, technologies, and processes to create human repairing or replacing cell, tissue, and organ products [5-17,20-27] (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8).
The clinical applications of hESC therapy derivatives as novel stem cell medicine for neural and cardiac repair are new therapeutic market opportunities. Unlike traditional drug development, novel neuronal or cardiac cellular therapeutic products of PluriXcel technology platforms are developed directly using human cells of neuronal or myocardium regenerative potential with proof-of-concept already established in humans, which simplifies the development process, lowers the costs, shortens the time consumption, and increases the probability of clinical success dramatically. PluriXcel therapeutic products - Xcel prototypes - have been developed specifically to address and overcome some major obstacles for successful pharmaceutical development of stem cell medicine, including offering the benefits in high efficiency, stability, low tumor risk, high purity, high efficacy in repair, as well as safety and large-scale production or manufacturing over all other existing approaches [7-17]. Such benefits of Xcel prototypes meet the commercial criteria in plasticity, specificity, and stability for designating any human stem cells as a cell therapy product for filing an IND application with the FDA and entry into human trials or first-in-human studies, thus will dramatically reduce the costs of registration pipeline as it reaches cGMP-compliance and clinical phase, and significantly increase the success rate in clinical trials [7-17].
Direct Conversion of Non-Functional Pluripotent hESC into Functional Somatic Elements Offers Scalable Platforms for Replacement Tissue and Organ Reconstitution
PluriXcel technology breakthrough offers a practical strategy to convert non-functional pluripotent cells directly into a specific functional lineage, which has built the platform for reconstitution or reconstruction of functional tissues and organs in a 3D setting that reflect the cellular characteristics and function of the in vivo human organ system to address unmet medical needs. The brain and the heart are among the first organs formed from the cells of the ICM or epiblast of the human blastocyst in early embryogenesis. The neural or cardiac elements derived from the embryo-originated hESC, the in vitro representation of the ICM or epiblast of the human blastocyst, resemble the neural or cardiac cells in human brain or heart development in embryogenesis; therefore, they have the intrinsic potential to form human CNS circuitry of the brain or myocardium of the heart in tissue-specific complex biomimetic culture systems. Indeed, the RA signaling at the pluripotent stage of hESC under the defined culture leads to a cascade of three sequential stages of CNS development: neuroectodermal induction, neuronal progenitor specification, and neuronal cell type/subtype generation and maintenance, resembling the process of neurogenesis in human embryonic development; whereas the NAM signaling at the pluripotent stage of hESC under the defined culture leads to a cascade of three sequential stages of cardiac development: cardiomesodermal induction, cardiac precursor specification, and cardiomyocyte generation and maintenance, resembling the process of cardiogenesis in human embryonic development [5-17,20-27] (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7 and Figure 8). In fact, the neuronal derivatives of embryo-originated hESC, following small molecule induction and suspension culturing, spontaneously progress and self-assemble into neuron circuitries reminiscent of the ventral mesencephalon of the human brain [5-12,20-25] (Figure 5). Similarly, the cardiomyocyte derivatives of embryo-originated hESC, following small molecule induction and suspension culturing, spontaneously progress and self-assemble into large beating cardiomyocyte clusters with strong rhythmic impulses reminiscent of the p-QRS-T-complexes seen from body surface electrodes in clinical electrocardiograms (ECG) [7-17,20,25-27] (Figure 7).
PluriXcel technology breakthroughs provide vital scalable human neuronal and cardiomyocyte cell sources for creation of CNS-or heart-related replacement tissue and organ products at a commercial or clinical scale necessary for CNS or heart tissue or organ replacement therapies. This hESC-based innovative regenerative medicine technology has built the platform for development of much-demanded reliable tissue-specific complex biomimetic culture systems for 3D CNS and myocardial tissue assembly or reconstitution from hESC neuronal and cardiomyocyte derivatives, respectively, providing advancements over current human stem- and progenitor-derived cell-type selectivity approaches through improvements in neuronal or cardiomyocyte differentiation efficiencies, microenvironment niches, multi-cellular 3D architecture, and cellular composition and function that represent the characteristics of the CNS or contractile heart muscles and demonstrate reproducible cellular signatures, circuitries, and functional outputs that can replicate aspects of the human CNS or heart. The development and utilization of multi-cellular 3D CNS or heart models that represent the 3D architecture and cellular composition and simulate functional characteristics of the human CNS or heart will pave the way for further development of cutting-edge automated high-content biofabrication systems for systematic functional assembly or reconstitution of the in vitro functional tissues or organs in a 3D setting that reflect the characteristics and function of the in vivo human organ system, enabling automated high content and high-throughput analysis and systems developmental biology modeling of the human CNS or heart circuitries and dynamics in order to understand the complex human development and disease pathologies of a wide range of CNS and heart disorders.
PluriXcel technology breakthroughs enable the well-controlled induction of hESC at the pluripotent stage directly and efficiently into CNS- or heart-related cells, shaping the future of medicine by providing novel, unique solutions at a clinical grade. Such advancements in stem cell medicine are expected to bring groundbreaking hESC-based technology platforms and innovative regenerative approaches to transform biomedical research. However, since initial derivation of hESC lines from the IVF leftover embryos in 1998, the social and legal controversies constantly surrounding hESC research have resulted in decade long policy battles concerning Federal funding for hESC research [5,7]. In 2009, President Obama lifted Bush administration's restriction on National Institutes of Health (NIH). As a result, the NIH registry has been expanded to include hundreds of hESC lines since. So far, those hESC lines on NIH registry are exclusively for research purpose only, but none of the hESC lines eligible for Federal funding are biologics-free clinical-grade hESC suitable for patient uses or clinical trials. Despite some progresses, Federal funding remains barred from any activities associated with the derivation of hESC from IVF leftover embryos. During the past two Presidential terms, all the efforts of Obama administration to relax policy and increase NIH funding for hESC research have been stalled by fierce moral oppositions and subsequent court battles, and Obama administration's relaxed policy favorable NIH funding for hESC research might face reversal by the incoming administration. In fact, most of the NIH stem cell research funding during the past two Presidential terms of Obama administration has ironically been allocated to the over-promised adult alternative iPS cells, which, despite well-funded, have generated neither intended public interests nor promised medicinal benefits to really push the stem cell medicine field forward. In spite of such uncertainty, future efforts to relax policy and allocate funding to support such hESC research for the development and application of innovative regenerative medicine technologies to improve our understanding of the dynamics and complexity of the human tissue and organ function in development and disorders are crucial in order to dramatically increase the overall turnover of investments in biomedical sciences to optimal treatment options or cures for a wide range of incurable or hitherto untreatable CNS and heart disorders.
References
- Thomson JA, Itskovitz-Eldor J, Shapiro SS, et al. (1998) Embryonic stem cell lines derived from human blastocysts. Science 282: 1145-1147.
- Benjamin E Reubinoff, Martin F Pera, ChuiYee Fong, et al. (2000) Embryonic stem cell lines from human blastocysts: somatic differentiation in vitro. Nature Biotechnology 18: 399-404.
- Cowan CA, Klimanskaya I, McMahon J, et al. (2004) Derivation of embryonic stem cell lines from human blastocysts. N Engl J Med 350: 1353-1356.
- Steiner D, Khaner H, Cohen M, et al. (2010) Derivation, propagation and controlled differentiation of human embryonic stem cells in suspension. Nat Biotechnol 28: 361-364.
- Parsons XH, Teng YD, Moore DA, et al. (2011) Patents on technologies of human tissue and organ regeneration from pluripotent human embryonic stem cells. Recent Pat Regen Med 1: 142-163.
- Parsons XH, Teng YD, Snyder EY (2009) Important precautions when deriving patient-specific neural elements from pluripotent cells. Cytotherapy 11: 815-824.
- Parsons XH (2016) PluriXcel: Emerging Technologies of Regenerative Medicine. LAP LAMBERT Academic Publishing.
- Xuejun H Parsons (2016) Human pluripotent stem cell-based PluriXcel technology platforms provide stem cell treatment: development and manufacturing innovations for progressing to the clinic. Journal of Regenerative Therapeutics 1: 1-8.
- Parsons XH (2015) Directing pluripotent human embryonic stem cells towards lineage-specific cell therapy derivatives for regenerative medicine. In: Nancy Smyth Templeton, Gene and Cell Therapy: Therapeutic Mechanisms and Strategies. (4th edn), CRC Press, Chapter 31: 795-818.
- Parsons XH (2013) Constraining the pluripotent fate of human embryonic stem cells for tissue engineering and cell therapy - the turning point of cell-based regenerative medicine. Br Biotechnol J 3: 424-457.
- Parsons XH (2013) Embedding the future of regenerative medicine into the open epigenomic landscape of pluripotent human embryonic stem cells. Ann Rev Res Biol 3: 323-349.
- Parsons XH (2013) Current state of regenerative medicine: moving stem cell research from animals into humans for clinical trials. JSM Regen Med Bio Eng 1: 1005.
- Parsons XH (2012) Mending the broken heart - Towards clinical application of human embryonic stem cell therapy derivatives. J Clinic Exp Cardiology 3: e116.
- Parsons XH (2013) Exploring future cardiovascular medicine: heart precursors directed from human embryonic stem cells for myocardium regeneration. Cardiol Pharmacol 2: e110.
- Parsons XH (2013) Cellular medicine for the heart - the pharmacologic utility and capacity of human cardiac stem cells. J Clinic Exp Cardiology S11: e001.
- Parsons XH (2013) Reviving cell-based regenerative medicine for heart reconstitution with efficiency in deriving cardiac elements from pluripotent human embryonic stem cells. Cardiol Pharmacol 2: e112.
- Parsons XH (2014) The designation of human cardiac stem cell therapy products for human trials. J Clin Trial Cardiol 1: 02.
- Parsons XH (2012) The dynamics of global chromatin remodeling are pivotal for tracking the normal pluripotency of human embryonic stem cells. Anat Physiol.
- Parsons XH (2014) The openness of pluripotent epigenome - defining the genomic integrity of stemness for regenerative medicine. Int J Cancer Ther Oncol 2: 020114.
- Parsons XH (2016) Direct conversion of pluripotent human embryonic stem cells under defined culture conditions into human neuronal or cardiomyocytes cell therapy derivatives. Methods Mol Biol 1307: 299-318.
- Parsons XH, Teng YD, Parsons JF, et al. (2011) Efficient derivation of human neuronal progenitors and neurons from pluripotent human embryonic stem cells with small molecule induction. J Vis Exp 56: e3273.
- Parsons XH (2013) Human stem cell derivatives retain more open epigenomic landscape when derived from pluripotent cells than from tissues. J Regen Med 1: 2.
- Parsons XH, Parsons JF, Moore DA (2012) Genome-scale mapping of microRNA signatures in human embryonic stem cell neurogenesis. Mol Med Ther 1: 2.
- Parsons XH (2012) An engraftable human embryonic stem cell neuronal lineage-specific derivative retains embryonic chromatin plasticity for scale-up CNS regeneration. J Reg Med Tissue Eng 1: 3.
- Parsons XH (2012) MicroRNA profiling reveals distinct mechanisms governing cardiac and neural lineage-specification of pluripotent human embryonic stem cells. J Stem Cell Res Ther 2: 124.
- Parsons JF, Smotrich DB, Gonzalez R, et al. (2012) Defining conditions for sustaining epiblast pluripotence enables direct induction of clinically-suitable human myocardial grafts from biologics-free human embryonic stem cells. J Clin Exp Cardiolog S9: 001.
- Parsons XH, Teng YD, Parsons JF, et al. (2011) Efficient derivation of human cardiac precursors and cardiomyocytes from pluripotent human embryonic stem cells with small molecule induction. J Vis Exp 57: e3274.
Corresponding Author
Xuejun H Parsons, San Diego Regenerative Medicine Institute & Xcelthera Inc, San Diego, CA, USA.
Copyright
© 2017 Parsons XH. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.