Colophospermum Mopane Leaf Production and Phenology in Southern Africa's Savanna Ecosystem - A Review
Abstract
colophospermum mopane, commonly known as mopane, is a dominant tree or shrub in the mopane woodland. It is widely distributed in the low-lying areas of southern Africa's savanna. Despite its wide distribution, ability to survive water stress conditions, and value as a source of browse for herbivores, especially during the dry season, there is limited knowledge on mopane leaf production and phenology as well as its implications to browsers. This knowledge gap is making it difficult for game ranchers and conservationists to fully understand the availability and quality of mopane browse in different seasons. As a result, this paper reviewed and integrated the available information on mopane leaf production and phenology in order to understand the amount and quality of mopane leaf forage during different seasons. This review shows that mopane leaf production and phenology are mainly triggered by rainfall. Mopane leaf flush starts after the first summer rains and reaches optimum productivity between February and March. The leaves begin to shed around May, following the onset of the dry season. Most of the dry or dead leaves remain on the tree until they are blown off by wind, especially in August. The leaves on the ground contribute significantly to the diet of browsers in mopane woodland during the dry season.
Keywords
colophospermum mopane, Phenology, Production, Leaf quality, Browsers, Southern africa
Introduction
colophospermum mopane (Benth.) J. Léonard, commonly known as mopane [1], is a dominant tree or shrub in the mopane woodland [2]. Mopane woodland is distributed in the low-lying areas of southern Africa's savannas, between latitudes 9°S and 25°S [2]. The total area covered by mopane woodland in its distribution in southern Africa is 555 000 km2 [3], which amounts to about 35% of the 1.5 million km2 of savanna in southern Africa [4]. Mopane is distributed in the hot and dry environments of southern Africa's savanna biome [5]. It is physiologically adapted to tolerate water stress and high temperature conditions [6]. Mopane folds its leaflets together [7], closes the stomata [8], and directs intercellular water to flow towards the inside of cells [9], particularly during the heat of the day [7]. These physiological responses enable mopane leaves to restrict water loss through transpiration [10], which allows mopane to tolerate drought conditions [6,11].
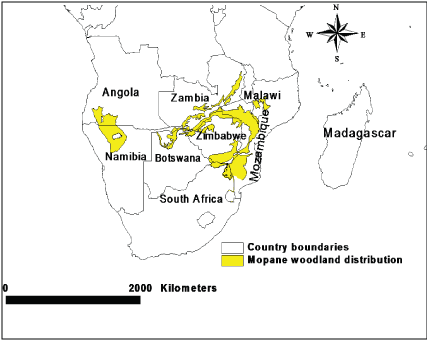
The distribution of mopane ranges from southern Angola and northern Namibia across Botswana and Zimbabwe to central and southern Mozambique, and from the Luangwa valley in Zambia and central Malawi to northern South Africa (Figure 1), with its southern-most distribution in the Klaserie Private Nature Reserve, south of the Olifants River [12]. Mopane is distributed in areas that have a variety of soil types, but mainly survives on alkaline soils [2], receiving low to moderate annual rainfall ranging from 400 mm to 800 mm [7], with mean daily temperatures ranging from 12 ℃ to 31 ℃ [13]. Acidic soil [10], a low winter temperature of < 5 ℃ and frost [14] are limiting factors for mopane distribution [15]. In addition, frequent fires can change the morphology of mopane from trees into shrubs [16].
Mopane is an irregular-shaped deciduous tree or shrub, which can grow up to 20 m high, with an erect narrow crown, although it can sometimes be found as an irregular deciduous 1-3 m high shrub [2]. The leaves of mopane are alternate, with a single pair of large triangular leaflets [10]. The two leaflets resemble the wings of a butterfly [17]. Young leaves are glossy and golden brown in colour, changing to green as they mature [18]. Most of the dry or dead leaves remain on the tree until they are blown-off by wind [19].
Mopane leaves are nutritious, containing between 6-13% of crude protein [20]. In addition, mopane maintains its foliage well into the dry season, thus providing important forage for browsers and intermediate feeders [21,22]. Dry mopane leaves are a valuable source of browse for herbivores [17] such as elephants (Loxodonta africana) [22], greater kudu (Tragelaphus strepsiceros) [23], and other ungulates found in mopane woodland, particularly during the dry season [24]. The nutritional value of mopane browse therefore explains why many rangelands, game reserves and national parks in the southern Africa's savanna biome are found in the mopane woodland [25].
Considering the ability of mopane to survive water stress conditions [6,11], and its value in supplying nutritious forage to browsers within its distribution range in southern Africa's savanna [17,26,27], knowledge of mopane leaf production and phenology becomes important. The key question is what is the amount and quality of mopane leaves during different seasons? This is an important question to answer because it has implications on browsers foraging behaviour, habitat selection as well as their spatial distribution in mopane woodland. The attainment of knowledge on mopane leaf availability and quality during different seasons will therefore require better understanding of leaf production and phenology. Unfortunately, limited research has been undertaken in these areas and the available information is old and scanty. Studies on mopane leaf production are very limited [28-32]. Limited studies on mopane leaf phenology have also been conducted [19,21,32]. Considering that mopane is widely distributed in southern Africa's savanna, this knowledge gap makes it difficult for game ranchers and conservationists to fully understand the availability and quality of forage in mopane-dominated areas. It further hinders effective monitoring of the state of health of mopane and the spatial distribution of browsers in the mopane woodland. The objective for this paper is to review and integrate the available and accessible information extensively in order to fully understand mopane leaf production and phenology as well as its implication for browsers in southern Africa's savanna. The availability of information is crucial in informing game ranchers and conservationists on how mopane woodland can best be managed in order to supply browsers with sustainable, adequate and quality browse. It is also critical for guiding future research and the management of mopane and the herbivores it supports through forage in the mopane woodland.
Methodology
Secondary method of data and information gathering was followed through extensive literature review from journal articles, books, academic thesis and dissertations, conference and workshop proceedings as well as from government, technical and scientific reports. The focus of the review was on mopane leaf production and phenology as well as its implications to browsers in the mopane woodland. The review primarily covers the distribution range for mopane woodland within the southern Africa's savanna. Data and information from literature was synthesised in order to determine the amount of mopane leaves and its phenology during different seasons. ArcView GIS 3.2 was used to produce the distribution map of mopane woodlands in southern Africa's savanna (Figure 1). The map was produced using Mucina and Rutherford [33] data on the vegetation of South Africa, Lesotho and Swaziland (VegMap), and White data on vegetation of Africa. In addition, based on the synthesis of review findings, a theoretic analysis of mopane leaf production and phenology was produced (Figure 2). Data and information gathered from the literature was further synthesised and integrated in order to determine the implication of mopane leaf production and phenology to browsers in the mopane woodland.
Mopane leaf production
It is important to assess mopane leaf production [29], because it has a significant effect on leaf availability to browsers [34]. However, the optimum productivity of mopane leaves is mostly affected by factors such as defoliation by Imbrasia belina [35], fires [16,36], browsing pressure [37,38], and climatic variability [30,36] including frost [14].
The effect of browsing on mopane leaf production was simulated through coppicing and pruning in the Limpopo Province, South Africa [30]. In addition, mopane trees were defoliated at 50%, 75%, 90% and 100% in the Kruger National Park, South Africa to simulate the effect of browsing on mopane leaf production [32]. The results showed that the number of leaves between different defoliation levels were not different [32]. The mean number of leaves per sapling remained highest for the control plants almost continuously. During early summer, the plants from the 100% defoliation treatment produced a larger numbers of leaves than those subjected to the 50% treatment. At the end of the experiment, the 100% defoliated plants had the smallest number of leaves [32], which suggests that heavy browsing could have a negative effect on mopane leaf production.
Defoliation of mopane leaves by mopane worms (Imbrasia belina) significantly reduced the density of mopane leaves as opposed to non-defoliated trees [28]. In addition, the number of mopane leaves on trees pruned by elephants (Loxodonta africana) was significantly lower than the number of leaves on non-pruned trees [28,31]. The causes of the reduction in the number and density of leaves in recovered defoliated and pruned mopane trees are uncertain, but it could be due to resources being stored for reproduction instead of regrowth of new leaves. However, pruning by elephants resulted in the production of longer shoots with larger leaves, while defoliation by mopane worms resulted in regrowth with shorter shoots and smaller leaves [28,31].
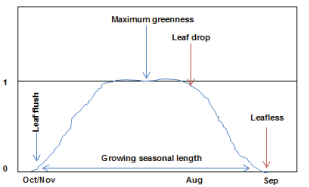
Other studies put forth that rainfall [30] and stem basal area [39] also have an influence on leaf production. This suggests that above average rainfall and trees with large basal stem areas could increase leaf production. However, mopane leaf production does not follow any consistent pattern during the growing season [30], but is greatly triggered by rainfall [34,40]. The production of mopane leaves starts after the first rain [18], normally during late October to early November [19,41]. The production of new leaves did not cease during the wet season, but continued to a lesser degree during the late wet season into the early dry season [23]. Mopane trees and shrubs reach optimum leaf production between February and March (Figure 2). The area of mature mopane leaflets ranges from 1507 mm2 to 2778 mm2 on average [30]. Growth season cumulative leaf production reaches its maximum in June and starts to decline in August (Figure 2), possibly because of a decline in soil moisture, following the decline in rainfall.
Plant thinning seems to increase mopane leaf biomass [34], but the quantity varies annually following rainfall patterns [40]. Estimates show that mopane leaf biomass in the northern Limpopo, South Africa is 1.1 tonnes/ha [42], which is lower than the 1.6 tonnes/ha estimated in south eastern Zimbabwe [43]. However, the total leaf dry matter of mopane in the Limpopo mopane woodland was 1.4 tonnes/ha [41], which is within the 0.6 tonnes/ha to 1.7 tonnes/ha estimated in the same locality [44]. An important point is that mopane contributes substantially to the total leaf dry matter in all communities in the Limpopo mopane woodland [41]. Therefore, it is the high leaf production of mopane [41] and its nutritional content [20] that makes it an essential species in providing forage for browsers, especially during the dry season in mopane-dominated areas.
Mopane leaf phenology
Phenology has been defined as the timing of biological events and their relationship to seasonal climatic changes [45]. Leaf phenology is thus defined in this review paper as the timing of leaf flush, growth and fall (Figure 2). Leaf flush refers to the production of new leaves, which grow until they shed or fall. Leaf flush and fall are attributed to changes in environmental conditions and also the effect of fire and browsers on the plant. Leaf phenology is thus an essential indicator of plant health and environmental stresses such as soil moisture, soil nutrients and solar radiation [46]. Phenology assists in monitoring and predicting vegetation responses to interannual climatic variability [4]. Changes in leaf phenology also indicate modifications in the environment. For instance, leaf flush indicates the availability of moisture in the soil, while leaf fall indicates a deficiency of soil moisture. Phenology is therefore recognised as one of the simplest and most effective indicators to study the impact of climate change [47,48], fires and herbivores on plants [49,50].
There are five main phenological stages in southern Africa's savannas, which are the pre-rain (September-October), early rainy (November-December), mid-rainy (January-February), late rainy (March-April) and dry seasons (May-August) [19]. The dry (May-September) and wet (October-April) seasons are critical period for determining leaf phenology and production. Mopane leaf flush occurs after the first summer rain [51], usually during October and November [7]. However, a delay in the first rain does not totally stop mopane from flushing its leaves. In countries such as Namibia and Botswana mopane leaf flush may be independent of rain [21]. It is suggested that the ability of mopane to flush its leaves before the first summer rain is mainly due to its ability to store energy, nutrients and water, which are used to flush new leaves even before the first rain.
Newly emerged mopane leaves are soft [52], appear at the beginning of November [41] and become leathery when they mature [52]. The mature green mopane leaves remain on the tree until they turn yellowish or reddish-brown with the onset of the dry season, particularly around May. However, most of the dry or dead leaves remain on the tree until they are blown off by wind [19], especially in August [41] to early September. Mopane leaves are shed in an irregular fashion at the onset of the dry season [51] and the trees are generally leafless from August to early October [41], though some of the leaves may remain on the tree until the beginning of summer. The leaves may be shed early in the dry season when rainfall is below average, but during above average rainfall, the leaves may remain on the plant until late into the dry season [53]. It thus appears that mopane leaf's phenological characteristics enable it to have a pro-longed growing season [46]. The growing season may last for 10 months (Figure 2), giving mopane longer leaf carriage than most other species in the mopane woodland.
The phenology of species in mopane woodland seems to be related to rainfall and temperature, but rainfall seems to be the main factor influencing variation in leaf phenology [54]. Studies in the savannas also confirm that soil moisture, as triggered by rainfall, is the major influence on the phenology of savanna species [55,56]. In addition to environmental influences on mopane leaf phenology, browsers such as mopane worms, greater kudu and elephants also have an influence on mopane leaf phenology. For instance, mopane worms browse 90% of mopane leaves [57,58], which disturbs leaf growth and phenology [35]. The leaves later regrow after defoliation [19], but leaf development and phenological stages are disturbed. The regrowth of leaves after trees have been defoliated could explain why mopane has a prolonged growing season, as illustrated in Figure 2.
Defoliation of mopane leaves by mopane worms seems to be a more significant disturbance of leaf phenology than browsing by mammalian herbivores. This is due to the high numbers of mopane worms on a tree, against the one or two browsing mammals, which feed on a few leaves and then move to the next tree. In addition, browsing of mopane leaves by mammalian herbivores during the growing season is limited by high production of secondary metabolites such as tannins and phenols [59]. This suggests that the impact on mopane phenological stages by browsing mammals could be minimal, but is significant for defoliation caused by mopane worms.
Implication of leaf production and phenology for browsers
Mopane leaf production and phenology have great implications for browsers because they determine the availability and quality of forage [60]. Browsers such as the greater kudu are affected by a shortage in forage supply [61], as well as the quality of the forage. The distribution and habitat selection of many browsers is therefore determined, among others, by forage availability [60] and quality in a particular season.
Seasonal differences in the relative contribution of woody plants to browsers' diet are also related to phenological changes during certain seasons and between seasons. However, seasonal variations in diet composition are not entirely attributable to variations in forage availability, but also to the accessibility of preferred species for browse [23]. Shortage of forage supply and quality can result in mortality of browsers such as the greater kudu, especially in fenced protected areas [34]. However, during the dry season (May to September), the average crude protein values in mopane leaves declined from 11.4% in May to 9.4% in September. The average concentration of phosphorus and calcium in mopane leaves declined from 0.20% and 2.01% in May to 0.12% and 1.38% in September, respectively [62]. Though nutritional content in mopane leaves is low during the dry season, high litter fall of mopane leaves during that season provides browsers with an alternative source of forage to survive [40] when forage is scarce, and most savanna species are leafless [22].
Leaf production therefore influences the time that browsers can stay in a particular habitat, while phenology influences the diet selection of browsers, as determined by forage quality, during different phenological stages. The selection of diet by browsers such as the greater kudu may vary between seasons, with species being favoured in one season but rejected in another [23]. For instance, mopane leaves are highly browsed during the dry season [27], but not preferred during the wet season, though the quality is higher [20], mainly because of a high concentration of secondary metabolites such as tannins and phenols [59].
During winter, fallen dry mopane leaves constitute a significant amount of leaves on the ground in the mopane woodland [41]. They thus add to a sustainable supply of forage to browsers in mopane-dominated areas and contribute immensely to the diet of browsers [27] such as the greater kudu [20,23], elephants [21,22,28] and other ungulates. High mopane leaf biomass on the ground during the dry season can therefore influence many browsers to survive in the mopane woodland during the dry season, as opposed to the wet season, because then there will be enough palatable forage in other areas. In addition, the ability of mopane to carry its leaves longer into the dry season [41] (Figure 2), when most savanna species are leafless [22], makes mopane the most influential fodder resource for many browsers in mopane woodland.
Conclusion
Leaf production and phenology are important parameters to measure the temporal and spatial changes in the leaves, which is critical in determining the availability and quality of forage for browsers in a particular season. It is also important in determining leaf physiology and plant stress. The availability and quality of leaves seems to be triggered by rainfall, while browsers disturb the growth and quality of leaves. Leaf production and phenology therefore play a significant role in determining the season in which mopane will be highly preferred by browsers. Mopane leaves are preferred by mammalian browsers during the dry season, when the leaves are dry, available on the ground, and secondary metabolites are relatively low. In addition, the high leaf production and the ability of mopane to carry its leaves longer into the dry season than most savanna species make it one of the important species in providing essential browse in the mopane woodland, and it is thus highly browsed during that period.
Acknowledgement
The authors thank the DAAD-NRF Doctoral Scholarship and the Limpopo Legislature for financial support.
References
- Léonard J (1949) Colophospermum. Notulae Systematicae IV (Caesalpiniaceae Amherstieae Africanae Americanaeque). Bulletin du Jardin Botanique de Létat (Bruxelles) 19: 388-408.
- Werger MJA, Coetzee BJ (1978) The Sudano-Zambezian region. In: Werger MJA, Biogeography and Ecology of Southern Africa. Dr. W. Junk, The Hague, 301-453.
- Mapaure I (1994) The distribution of colophospermum mopane (Leguminosae-Caesalpinioideae) in Africa. Kirkia 15: 1-5.
- White F (1983) The Vegetation of Africa. United Nations Educational, Scientific and Cultural Organization, Paris.
- Siebert F (2012) A phytosociological synthesis of mopaneveld vegetation at different spatial scales using various classification methods [thesis]. North-West University, Mahikeng.
- Choinski JS, Tuohy JM (1991) Effect of water potential and temperature on the germination of four species of African savanna trees. Annals of Botany 68: 227-233.
- Madams RW (1990) The biogeography of colophospermum mopane (Kirk ex Benth.) Kirk ex J. Léon. at its distribution limit in eastern Botswana [thesis]. Queen Mary Westfields College, University of London, London.
- Mantlana KB (2002) What determine the structure of colophospermum mopane under field conditions in North-Western Botswana. MSc Dissertation. University of Natal, Durban.
- Chen H, Jianga J (2010) Osmotic adjustment and plant adaptation to environmental changes related to drought and salinity. Environmental Reviews 18: 309-319.
- Henning AC (1976) A Study of edaphic factors influencing the growth of colophospermum mopane (Kirk ex Benth.) Kirk ex J. Léonard [thesis]. University of the Witwatersrand, Johannesburg.
- Johnson JM, Pritchard J, Gorham J, et al. (1996) Growth, water relations and solute accumulation in osmotically stressed seedlings of the tropical tree colophospermum mopane. Tree Physiol 16: 713-718.
- Witkowski ETF, O'Connor TG (1996) Topo-edaphic, floristic and physiognomic gradients of woody plants in a semi-arid African savanna woodland. Plant Ecology 124: 9-23.
- Makhado RA, Mapaure I, Potgieter MJ, et al. (2014) Factors influencing the adaptation and distribution of colophospermum mopane in southern Africa's mopane savannas - A review. Bothalia 44.
- Whitecross MA, Archibald S, Witkowski ETF (2012) Do freeze events create a demographic bottleneck for colophospermum mopane? South African Journal of Botany 83: 9-18.
- Stevens N, Swemmer AM, Ezzy L, et al. (2014) Investigating potential determinants of the distribution limits of a savanna woody plant: colophospermum mopane. Journal of Vegetation Science 25: 363-373.
- Gandiwa E, Kativu S (2009) Influence of fire frequency on colophospermum mopane and Combretum apiculatum woodland structure and composition in northern Gonarezhou National Park, Zimbabwe. Koedoe 51: 36-48.
- Timberlake JR (1995) colophospermum mopane. Annotated Bibliography and Review. The Zimbabwe Bulletin of Forestry Research No. 11. Forestry Commission of Zimbabwe, Bulawayo.
- Sebego RJG (1999) The ecology and distribution limits of colophospermum mopane in southern Africa. Botswana Notes and Records 31: 53-72.
- Guy P, Mahlangu Z, Charidza H (1979) Phenology of some trees and shrubs in Sengwa wildlife research area, Zimbabwe. South African Journal of Wildlife Research 9: 47-54.
- Hooimeijer JP, Jansen FA, de Boer WF, et al. (2005) The diet of kudus in a mopane dominated area, South Africa. Koedoe 48: 93-102.
- Styles CV, Skinner JD (2000) The influence of large mammalian herbivores on growth form and utilization of mopane trees, colophospermum mopane in Botswana's Northern Tuli Game Reserve. African Journal of Ecology 38: 95-101.
- Kos M, Hoetmer AJ, Pretorius Y, et al. (2012) Seasonal diet changes in elephant and impala in mopane woodland. European Journal of Wildlife Research 58: 279-287.
- Curlewis BJ (2014) The seasonal feeding composition of kudu (Tragelaphus strepsiceros) and movement of kudu, eland and nyala in the mopani veld of the Limpopo Province [thesis]. University of Limpopo, Sovenga.
- Makhado RA, Potgieter MJ, Luus-Powell WJ, et al. (2015) Tragelaphus strepsiceros browse during the dry season in the mopani veld of Limpopo Province, South Africa. Transaction of the Royal Society of South Africa 71: 17-21.
- Bonsma JC, Du Toit JD (2010) Game Ranch Management. (5th edn), Van Schaik Publishers, Pretoria.
- Mosimanyana BM, Kiflewahid B (1988) Value of browse as ruminant feed: The case of colophospermum mopane. IDRC Digital Library, Gaborone.
- Macala J, Sebolai B, Majinda RR (1992) colophospermum mopane browse plant and sorghum stover as feed resources for ruminant during the dry season in Botswana. In: Stares JES, Said AN, Kategile JA, The Complementarity of Feed Resources for Animal Production in Africa. Proceedings of the Joint Feed Resources Networks Workshop held in Gaborone, Botswana, 4-8 March 1991. African Feeds Research Network. International Livestock Centre for Africa, Addis Ababa, Ethiopia.
- Hrabar HDNK (2005) Indirect interactions between elephants (Loxodonta africana) and mopane caterpillars (Imbresia belina) through their shared food resource - mopane trees (colophospermum mopane) [thesis]. University of Pretoria, Pretoria.
- Ghazoul J (2006) Mopane Woodlands and the Mopane Worm: Enhancing rural livelihoods and resource sustainability. DFID Project Reference Number R 7822, 1-6.
- Wessels D, Mushongohande M, Potgieter M (2006) Mopane tree ecology and management. In: Ghazoul J, Mopane Woodlands and the Mopane Worm: Enhancing Rural Livelihoods and Resource Sustainability. DFID Project Reference Number R 7822, 7-17.
- Hrabar H, Hattas D, Du Toit JT (2009) Differential effects of defoliation by mopane caterpillars and pruning by African elephants on the regrowth of colophospermum mopane foliage. Journal of Tropical Ecology 25: 301-309.
- Kohi EM, de Boer WF, Slot M, et al. (2010) Effects of simulated browsing on growth and leaf chemical properties in colophospermum mopane saplings. African Journal of Ecology 48: 190-196.
- Mucina L, Rutherford MC (2006) The vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. South African National Biodiversity Institute, Pretoria.
- Smit GN (2001) The influence of tree thinning on the vegetative growth and browse production of colophospermum mopane. South African Journal of Wildlife Research 31: 99-114.
- Adelabu AS (2013) The remote sensing of insect defoliation in mopane woodland [thesis]. College of Agriculture, Engineering and Science, University of KwaZulu-Natal, Pietermaritzburg, South Africa.
- Seydack AHW, Vermeulen WJ, Knox N, et al. (2014) Forage quality for grazing herbivores in a semi-arid savanna system as affected by fire and rainfall (Kruger National Park). Scientific Report Number 03/2014, South African National Parks, Skukuza.
- Lewis DM (1991) Observations on tree growth, woodland structure and elephant damage on colophospermum mopane in Luangwa Valley, Zambia. African Journal of Ecology 29: 207-221.
- Ben-Shahar R (1993) Patterns of elephant damage to vegetation in northern Botswana. Biological Conservation 65: 249-256.
- Scholes RJ (1990) The regrowth of colophospermum mopane following clearing. Journal of Grassland Society of Southern Africa 7: 147-151.
- Mlambo D, Nyathi P (2008) Litterfall and nutrient return in a semi-arid southern African savanna woodland dominated by colophospermum mopane. Plant Ecology 196: 101-110.
- Dekker B, Smit GN (1996) Browse production and leaf phenology of some trees and shrubs in different colophospermum mopane savanna communities. African Journal of Range and Forest Science 13: 15-23.
- Smit GN (1994) The influence of intensity of tree thinning on mopani veld [thesis]. University of Pretoria, Pretoria.
- Mutakela PS (2009) Biomass prediction models for colophospermum mopane (mopane) in Botswana. University of Stellenbosch, Stellenbosch.
- Smit GN, Rethman NFG (1998) Root biomass, depth distribution and relationships with leaf biomass of colophospermum mopane. South African Journal of Botany 64: 38-43.
- Lieth H (1974) Purposes of a phenology book. In: Lieth H, Phenology and Seasonality Modeling. Springer-Verlag, New York, 3-19.
- Fuller DO (1999) Canopy phenology of some mopane and miombo woodlands in eastern Zambia. Global Ecology and Biogeography 8: 199-209.
- Menzel A (2002) Phenology: Its importance to the global change community. Climatic Change 54: 379-385.
- IPCC (2007) Climate Change 2007: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge.
- Scholes RJ, Archer SR (1997) Tree-grass interactions in savannas. Annual Review of Ecology and Systematics 28: 517-544.
- Sankaran M, Hanan NP, Scholes RJ, et al. (2005) Determinants of woody cover in African Savanna. Nature 438: 846-849.
- http://r4d.dfid.gov.uk/PDF/Outputs/Forestry/R7822_-_Q2_-_Management_mopane_veld.pdf.
- Potgieter MJ, Wessels DCJ (1998) The anatomy and morphology of petioles and leaflets of Hardwickia mopane (Leguminosae: Caesalpinioideae). South African Journal of Botany 64: 281-289.
- Chidumayo E, Frost P (1996) Population biology of miombo trees. In: Campbell B, The Miombo in Transition: Woodlands and Welfare in Africa. Center for International Forestry Research, Indonesia, 59-72.
- Chidumayo EN (2001) Climate and phenology of savanna vegetation in southern Africa. Journal of Vegetation Science 12: 347-354.
- Archibald S, Scholes RJ (2007) Leaf green-up in a semi-arid African savanna - separating tree and grass responses to environmental cues. Journal of Vegetation Science 18: 583-594.
- Wessels K, Steenkamp K, von Maltitz G, et al. (2011) Remotely sensed vegetation phenology for describing and predicting the biomes of South Africa. Applied Vegetation Science 14: 49-66.
- Ditlhogo M, Allotey J, Mpuchane S, et al. (1996) Interactions between the mopane caterpillar, Imbrasia belina, and its host, colophospermum mopane in Botswana. In: Flower C, Wardell-Johnson G, Jamieson A, Management of Mopane in Southern Africa. Proceedings of the workshop held at Ogongo Agricultural College, Northern Namibia, 26-29 November 1996. University of Namibia, Windhoek, 45-48.
- Toms R, Thagwana M (2005) On the trail of missing mopane worms. Science in Africa.
- Wessels DCJ, van Der Waal C, de Boer WF (2007) Induced chemical defences in colophospermum mopane trees. African Journal of Range and Forage Science 24: 141-147.
- Janecke BB, Smit GN (2011) Phenology of woody plants in riverine thicket and its impact on browse availability to game species. African Journal of Range and Forage Science 28: 139-148.
- Owen-Smith N (1979) Assessing the forage efficiency of a large herbivore, the kudu. South African Journal of Wildlife Research 9: 102-110.
- Makhado RA, Potgieter MJ, Luus-Powell WJ (2016) Nutritional value of colophospermum mopane as source of browse and its chemical defences against browsers: A review. Journal of Animal & Plant Sciences 26: 569-576.
Corresponding Author
Rudzani A Makhado, Department of Biodiversity, University of Limpopo, P/Bag X1106, Sovenga, South Africa.
Copyright
© 2018 Makhado RA, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.