Soils at the Altitudinal and Northern Treeline: European Alps, Northern Europe, Rocky Mountains - A Review
Abstract
No treeline-specific soil types exist. The treeline ecotone is usually characterized by a mosaic of soil types closely related to the locally varying conditions. Microtopography and the patchiness of the plant cover play an important role in this respect. Most soils in the treeline are shallow, and rich in skeletal material. Freeze-thaw processes are important agents for pedogenesis. Natural disturbances such as landslides, avalanches and fire affect soil development in the treeline ecotone. Human impact has also been an important factor historically. Fossorial mammals and wild-living and domestic ungulates have been equally-involved in soil development. In the long-term, forest expansion into currently treeless terrain within the treeline ecotone and beyond the existing tree limit will be closely associated with changes in soil conditions irrespective of whether it is driven by climate warming and/or cease of pastoral use.
Keywords
Mountains, Pedogenesis, Soil mosaic, Soil types
Introduction
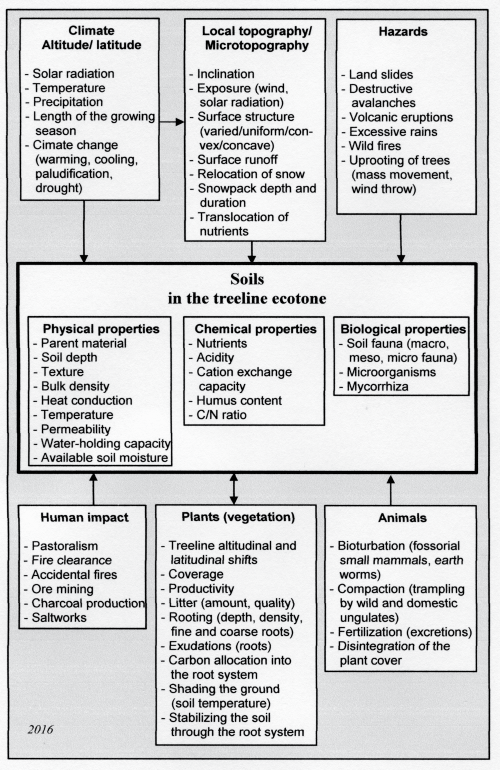
Treeline position, spatial pattern and dynamics depend on a multitude of interacting factors [1-4]. Among them soil is undoubtedly one of the most complex factors (Figure 1). However, the influence of soil on the treeline ecotone is usually less conspicuous than the effects of other factors such as heat deficiency, wind, and snowpack on tree growth and treeline spatial patterns. Thus, in most treeline studies, soils have been considered rather cursorily.
In the European Alps, research on soils in the treeline ecotone and in the alpine zone has been stimulated already almost 50 years ago by the challenge of restoring high elevation forests and afforestation above the present forest limits [5-9]. Broad information is also available on alpine soils in the Rocky Mountains [10-19]. Recently high-elevation soils in a worldwide view are considered in a FAO report and with focus on Asia by Müller, et al. [20,21]. In addition, Egli, et al. presents a review on soils of mountainous landscapes [22].
The possible influence of global warming on soil ecological conditions, especially in cold regions, has increasingly attracted researchers' interest [19,20,23-29]. In high-mountains, soils, in particular, play an important role in maintaining ecosystem services [20,30], such as buffering surface runoff, flood control, preventing slope erosion, and reduction of slumps and landslides.
The objective of this paper is to highlight the multiple interactions of soil conditions and tree stands and their possible effects on treeline spatial pattern and dynamics in the European Alps, in northernmost Europe, and in the Rocky Mountains, where the authors studied treeline ecological conditions during many field campaigns (see reference list). In the Alps, most studies were carried out in the Engadine and its neighboring valleys (Switzerland). In the north, the authors focused on treeline patterns and dynamics in northernmost Finnish Lapland. In addition, the authors studied the treeline ecotone from the southern to the northern Rocky Mountains and in some other mountain ranges in the American west. We apply the term 'treeline' to the transition zone (ecotone) between closed subalpine or northern forests to the elevational or northern limit of tree species. Depending on the local conditions, the transition zone varies in width [31,32].
General Characteristics of Soils at Treeline
The relative young age of soils is a common characteristic of formerly glaciated mountain landscapes. Most soils in the treeline ecotone are shallow und rich in skeletal material. Block-rich saprolite is usually lacking a continuous soil cover. On mountain ridges that were not covered by Pleistocene glaciers, such as some broad, rolling high interfluves in the Colorado Rocky Mountains and in Glacier National Park (Montana), for example, deep soils that developed over longer temporal scales can be found on gentle terrain [14,33-35]. In the Alps, buried relict soils that developed during the last interglacial period [28], can be found in the treeline ecotone and in the alpine zone.
Influence of elevation
Soils in the treeline ecotone are influenced by the altitudinal gradients of temperature and precipitation (Figure 1). Heat deficiency is of overall importance. Changes in plant nutrients, for example, driven by decrease of temperature with elevation are associated with changes in soil organic matter and quality (C/N-ratio) [29]. As most soils in the treeline ecotone are shallow and rich in skeletal material, their water-holding capacity may be relatively low [35,36]. Soil moisture conditions may however vary considerably under the influence of local topography. Freeze-thaw processes (e.g., cryoturbation, frost heave, needle ice formation) are important factors for soil development [37-41].
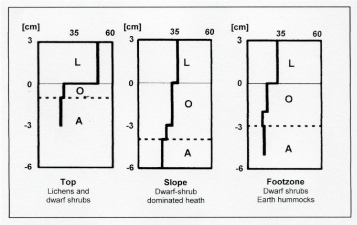
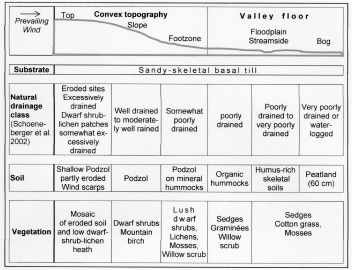
Very often, the soils in the treeline ecotone and in the lower alpine zone are relatively rich in humus due to slow decomposition [42]. In many cases, in addition to low soil temperatures and insufficient or excessive soil moisture, the wide C/N ratio of dwarf shrub and conifer litter and low pH values reduce microbial activity and decomposition [8]. In the treeline ecotone in the Upper Engadine (Switzerland), for example, C/N varies from 40-50 in dwarf shrub litter, whereas it is 25 in the litter of grass vegetation on pastures [43,44]. At treelines in northern Finland the authors found C/N varying from 50 in the litter layer (0-3 cm depth) on convex topography, covered with lichens and low dwarf shrubs, to 35 on the slopes, and 30 in the foot zone (dwarf shrubs, podzolised mineral hummocks). In the organic layer and A horizons C/N was ≤ 20 on top and in the foot zone, while ranging between 20 and 32 on the slopes of convexities (Figure 2) [45]. Podzolisation in and above the treeline ecotone is reduced due to low plant productivity and high evaporation [43,44]. High evaporation loss is a result of patchy plant cover, permanent wind and - at high elevation - intense solar radiation and relatively low air pressure. Low nitrogen availability and phosphorus deficiency are also characteristic of the altitudinal and northern treelines [16,18,46-48]. Moreover, the decrease of ectomycorrhizal fungal taxa is at least partly an elevational effect [48-51].
Despite the general elevational effects on soil formation, there are no treeline-specific soil types in the temperate mountains. Instead, a mosaic of soil types prevails, closely related to the locally varying conditions, among which local topography is the dominant factor. The overlapping influences of parent material together with soil properties and plant cover may be more important than elevational effects [5].
Disturbances
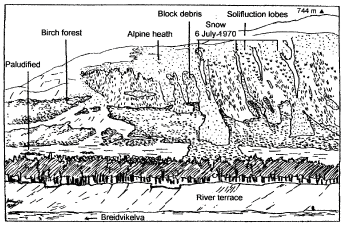
Disturbances by landslides, soil creep, and avalanches not only destroy trees but also affect soils in the treeline ecotone (Figure 1). Uprooting of trees, for example, by avalanches may locally initiate soil erosion. On steep slopes, soil development was often interrupted through downslope redistribution of soil material. Thus, truncated or buried soils are very common. Large block fields often cause a low elevation of the actual treeline on steep terrain. On unstable slope debris, soil formation usually remains at an initial stage for a long time. However, if coarse block-rich debris in backslope and footslope positions becomes covered with finer enhanced water-holding material from the upper slope soil formation will accelerate. In northern Norway, for example, Holtmeier [52] observed steep block debris areas partially covered with relatively fine material from solifluction lobes that had collapsed when entering the steep trough wall from above (Figure 3). The accumulated lobe material increases the water holding capacity on the otherwise extremely dry block debris and thus facilitates the establishment of birches, and willow shrub and other vegetation. Their root system stabilize the soil on steep terrain. Similar observations have been reported from steep mountain terrain in the Yukon Territory [53].
On steep alpine pastures, turf exfoliation [54,55] may truncate the soil profile and expose the mineral soil. Turf exfoliation is often triggered by cattle or wild ungulates that destroy the plant cover by trampling (Figure 4). Once the mineral soil is exposed, needle ice formation may enforce erosion by wind and surface runoff. On many subalpine and alpine pastures that have been grazed for hundreds of years, a dense network of cattle tracks has developed, which often enforces linear erosion through surface runoff.
During the Holocene, climatically-driven and anthropogenically influenced treeline fluctuations have influenced soils at regionally and locally varying intensities [31]. Pastoral use, forest fires, ore mining, charcoal production, and salt works played a major role in many treeline areas. In the Alps, for example, human impact has lowered treeline by 150 m to 300 m compared to its uppermost position during the postglacial optimum. In many areas, human impact dates back several thousand years [31,56-60]. The practice of fire clearance of high-elevation forests has been used for the past 4,500 years to create alpine pastures. In some areas, fire clearing started even earlier, probably during the Neolithic [61,62]. In the Himalayas, for example, a depression of more than 500 m is likely, mainly due to pastoral use [63]. In the Rocky Mountains, the oldest artifacts found in the treeline ecotone are over 8,000-years-old [64]. Thus, it is possible the first local human impact on the treeline occurred long before the Europeans arrived and caused severe disturbances, often combined with the use of fire. Fires often destroy the organic layers for the most part and locally expose the ground to splash, sheet, rill and gully erosion and/or strong winds in the burn areas (Figure 5). The loss of the organic layer resulted in nutrient impoverishment and reduced water holding capacity. Hence, soil susceptibility to erosion increased.
On heavily grazed pastures in the subalpine and alpine zone, mycorrhiza declined locally due to soil compaction and resultant stagnant water conditions. Moreover, the abundance of mycorrhizae also decreased in sites where permeable substrate, high soil temperatures or wind cause dry conditions [49]. In northern Norway and Finland, overgrazing by semi-domestic reindeer had similar effects on soil erosion in the treeline ecotone and above [45,52,65-67], Ectomycorrhiza make organically bound soil nutrients available to the plants and contribute to rapid turnover of soil organic carbon stock in the organic horizons [68].
Wildfires have locally affected soils in the treeline ecotone. At the northern treeline, destruction of the forest by fire-initiated paludification has led to a thicker moss cover, and accumulation of dead organic matter (Histosols) [69]. In areas of thawing permafrost and increased paludification, the forest has been regionally retreating to a more southern position [70].
Not least, fossorial animals (e.g., ground squirrels, pocket gophers, marmots) as well as wild-living ungulates locally influenced soil development (bioturbation, nutrient fluxes, erosion) (Figure 1). Animals' influence on soils in the treeline ecotone has widely been disregarded, with a few exceptions [31,52,71-73].
Variety of Soils in the Treeline Ecotone
A great variety of soils can be found in the treeline ecotone: Shallow and deep soils, truncated and buried soil profiles, soils rich or poor in nutrients, water-logged and well-drained soils [74]. In the treeline ecotone and alpine zone, both well-watered or adequately drained sites are usually less common compared to sites with insufficient or excessive moisture. Depending on substrate chemistry, substrate permeability and age of soil formation, Leptosols or Regosols developed. Plant cover varies with pH. On silicate substrate, the spectrum of soils in the treeline ecotone and lower alpine zone ranges from initial soils to fully developed Podzols and Cambisols, usually covered with zone typical acidic dwarf shrub vegetation and Carex curvula swards.
Locally varying soil moisture, as result of the direct influence of local topography (e.g., runoff) and wind-mediated relocation of winter snow (Figure 1), decomposition, nutrient content, nutrient availability, soil temperature (particularly near the soil surface), bulk density, heat conductance, and heat capacity are important for soil development, seedling establishment and performance of young growth in and above the treeline ecotone. Long-lasting low soil temperatures reduce mineralization and nutrient availability while simultaneously affecting photosynthesis, carbon investment, and thus root growth and nutrient uptake [18,75,76]. Available soil moisture plays a major role in this respect. However, under well insulating deep snowpack, microbial activity and decomposition continue significantly even outside the growing season [77-81].
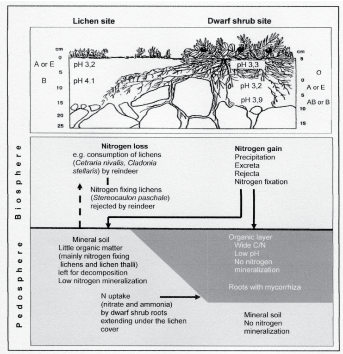
While soils, particularly their organic layers, are the main nutrient source for plants, the latter influence pedogenesis by carbon allocation, nutrient uptake, exudations from the roots, amount and quality of the litter (e.g., nitrogen content, retention time of nutrients, acidity) all of which serve to modify the microsites [7,82,83]. In the treeline ecotone in Lapland, for example, the nitrogen cycle in microsites covered by dwarf shrubs was found to be different from that of lichen-covered sites (Figure 6) [46].
In the treeline ecotone, present vegetation patterns reflect, to a certain extent, the multiple interactions of above-ground and below-ground processes. On crystalline parent material in the Central Alps for example, deep Podzols are associated with dwarf shrub-conifer forests. Thus, occurrences of well-developed Podzols above the present forest have been considered indicators of former forests [5,6,84,85] that were previously destroyed by humans. The Podzols probably developed during the last interglacial period [28]. Although well differentiated Podzol profiles were not always found, podzolisation could be evidenced analytically [86]. Podzolisation, however, may also occur under dwarf-shrub cover above the current treeline [44,86]. In the Alpine zone, percolating melt water may have a greater effect on podzolisation than vegetation [29]. On carefully managed alpine pastures, Cambisols have frequently developed. Soon after abandonment, however, dwarf shrubs recolonized these pastures and initiated podzolisation [87] when sufficient percolating water was present.
On well-drained terrain in the treeline ecotones of northern Europe, only shallow Podzols developed due to low productivity of the oligotrophic dwarf-shrub-lichen heath [88] and reduced litter supply. In addition, seasonally frozen ground prevents effective leaching. Under clusters of trees, more organic matter usually accumulates. In addition, open birch stands trap blowing snow which may increase the amount of melt water that would translocate minerals and dissolved substances into the soil. On the rolling land surfaces and low fells (< 1,000 m), annual precipitation at the treeline is less in comparison to the subalpine/alpine zone in the Alps.
Like relict Podzols in the Alps, 'subalpine Podzols' [89] have been considered as a relict of a warmer period (5.000-3.000 BP), when the treeline was 150-200 m higher than its present elevation [90]. In the Central Swedish Scandes, relict Podzols [91] together with mega fossils indicate the presence of conifers (forest?) at about 300-400 m above the current treeline [92]. On wind-swept and well-drained convex topography, the authors [93] found Podzol remains under partly eroded peat layers that probably accumulated during a wet and cool climate (since about 1,000 yr BP and 'Little Ice Age'), and under the influence of good drainage conditions, most likely under mountain birch stands with dwarf shrub under-growth. However, in sites, where dwarf shrub-lichen-heath on permeable substrate ensures low pH values, podzolization can also be evidenced at present [45,94]. Very shallow Podzols (formerly called 'Nanopodzols') on well-drained and wind-swept terrain, currently covered with low mats of dwarf shrub-lichen heath, are also common in other areas of northern Fennoscandia where permeable glacial till covers acidic bedrock [95].
On well-drained subalpine and alpine grassland in the Rocky Mountain treeline ecotone (Colorado Front Range) Cambisols prevail, whereas Podzols developed under coniferous forest [12,14]. In the upper part of the treeline ecotone (3,350-3,500 m), remnants of Podzols that had developed under downwind 'migrating' clonal conifer groups were observed [33,34]. In general, there is only a short period after snowmelt with above freezing temperatures and sufficient percolation to ensure the essential pre-conditions for leaching.
Soil formation does not just depend on local factors but may also be influenced by allochthonous impacts such as deposition of eolian dust carried great distances by prevailing winds, exhibited for example by the 'alpine loess' in the Colorado Front Range [13,15,33,34,96], or Saharan dust in the Alps [97-100] and dust from inland deserts (Taklimakan; Gobi) in the Tien Shan and Altai ranges [101]. In the Rocky Mountains, dust deposition increased considerably after the intensification of grazing and agriculture had caused widespread disturbance of soils on the Colorado Plateau and other desert regions of the western United States during the 1880s. In addition, recurrent periods of severe drought regularly increase emission of dust from the desert areas and deposition in the Rocky Mountains [102,103]. In the windy treeline ecotone on the Front Range, the authors found enriched silty eolian material in the soil under and downwind of tree islands, i.e. at sites accumulating blowing snow in winter [33,34]. The eolian dust is rich in calcium, which buffers soil acidity and increases cation exchange capacity. Thus, decomposition may accelerate and nutrient availability increase. Highest Ca contents and corresponding high pH values and CEC were measured at the leeward sites, while under the tree islands pH values were relatively low and initial podzolisation occurred [33]. The relatively high amounts of fine material that were found in the topsoil of very shallow Podzols in the treeline ecotone in northern Norway have also been ascribed to deposition of eolian dust [104]. Thin, fine-sandy and silty layers that were found in organic hummocks in the treeline ecotone in northern Finland are also of eolian origin [45].
Topographic Control of Soil Distribution Pattern
While the distribution pattern of soil types in the treeline ecotone is only partly related to the altitudinal gradient of temperature and precipitation, pronounced differences can develop in response to local topography [7,8,14, 31,43,45,86,104-110]. The direct and indirect effects of topographical factors on soil forming processes and vegetation development are key factors in the treeline ecotone and the adjacent alpine zone. Local topography influences microclimates, snow relocation, depth and duration of the winter snowpack, distribution of soil temperatures, freeze-thaw cycles, soil moisture, and translocation of solid and dissolved substances by run-off, seepage and leaching (Figure 1) [31,108,111,112]. Pronounced differences exist between convex and concave sites and between northern and southern exposures. The same holds true for microbial biomass and humus forms [105,113].
Convexities
Soils on convexities are usually of coarse texture and rich in skeletal material due to surface runoff and/or prevailing strong winds. Frequently the organic layers and the upper mineral horizon on the top and upwind side of convex terrain are eroded. Needle ice formation was also frequently noted in such situations, sufficient fine sandy-silty material provided. Freeze-thaw of the topsoil and frost heaving may cause mechanical damage of fine roots of tree seedlings. Frost heave largely depends on the presence of fine soil texture. On the other hand, tree islands that became established on convex topography may accumulate snow that prevents the soil from freezing to great depth [33,112].
In many places, trampling effects by wild ungulates triggered or enforced soil erosion. In the mountain-birch treeline ecotone in northern Finnish Lapland, for example, the original acidic dwarf shrub-lichen heath on wind-swept convex terrain became fragmented or destroyed completely by excessive numbers of semi-domestic reindeer (Figure 7). Under such conditions up to 60-90% of the soil profile can be eroded. Occasionally only down-wind 'migrating' wind scarps (Figure 8) were left [34,114]. Skeletal material relatively increased on the surface, as wind removed the fine material. Organic carbon and total nitrogen content are low in such places, whereas pH (very strongly acid) is a little higher on the eroded sites than under dwarf shrub-lichen patches. Deep freezing in winter due to missing or episodic winter-snow cover, frequent dry conditions (soil moisture often < 20% volume) and lack of nutrients can impede establishment of mountain birch seedlings in such places [115]. Warming climate may exacerbate drought stress that would affect not only the above-ground tissues of the seedlings and young trees but also adversely affect their fine roots in the organic layer and topsoil.
In high mountains, such as the American Rockies and the central European Alps, overheating may locally occur at sun-exposed but wind-protected sides of ribs and other convexities in the treeline ecotone, under dry and calm weather conditions on and immediately below dark and dry soil surfaces that are either sparsely vegetated or not vegetated at all. Maximum absolute temperatures may range between 40-60 ℃ for several hours and for short time even higher [116,117]. Such conditions may be critical to fine roots near the soil surface, that are vulnerable to both deep freezing and occasional extremely high temperatures [115,118-121]. Such temperatures may also cause serious damage to tree seedlings through heat girdling [122]. Drought damage, however, is usually more common in such sites, particularly on rapidly draining soils. At greater depth, excessive low or high temperatures and damage to the roots do not usually occur.
In water-saturated substrates on sloping terrain, solifluction may affect tree seedlings. At similar elevations, freeze-thaw events are more frequent in the upper soil layers on sunny exposures than on shaded rib sides. By contrast, late-lying deep snowpack on the shaded side of convex micro-topography usually prevents soil freezing at greater depths [31,123-125], provided snow comes early in winter when the soil is still unfrozen. In spring, however, warming of the soil on the shaded and often snow-rich side of convexities is delayed due infiltration of cold melt water and evaporation from the wet surface. As a result, soil usually remains relatively cool until snow has disappeared [126-128] as was also observed at the downwind edge of compact conifer groups where large snowdrifts had built up [33,34,112,129]. At late thaw sites, soil temperatures may rise rapidly after the snowpack melted and soil dried [130]. Winter snowpack and its influence of soil temperature vary by slope, aspect, and climatic regions (maritime, continental).
The critical root zone temperature for root growth has been found to be about 6 ℃, which is near the worldwide mean soil temperature at climatic treeline [131]. Anyway, root growth in young conifers (Pinus cembra, Picea abies, Pinus mugo) at treeline (2,000 m-2,300 m) on Stillberg (near Davos, Switzerland) started shortly after snow melt at soil temperatures between 2℃ and 3℃ and ceased at the same temperatures in mid-October [132]. At late thaw sites soil temperatures rose rapidly. Shoot elongation began earlier than root growth [130]. Significant growth of young trees may however be delayed. Conditions will be different if significant amounts of blowing snow accumulate on the sun-exposed side, i.e. prevailing winds from directions opposite to direct solar radiation. In continental climates (e.g., central Rocky Mountains), melt water supply from late-lying snowdrifts on southern exposures may reduce drought stress and prevent overheating of the topsoil at the beginning of the growing season, thus facilitating potential tree seedlings [32].
In general, relatively dry conditions prevail on convex topography due to run-off, seepage, low water holding capacity (coarse texture, thin or lacking organic layer) and wind (Figure 9). Direct wind impact, soil freezing to great depth, and drought stress may impair seedling establishment. Moreover, wind (turbulent flow and evaporation) lowers soil temperature during the growing season and prevents overheating. In addition, wind relocates loose fine mineral material and organic matter from the top of convexities to their leeward slopes and adjacent depressions, where it partly accumulates. Moreover, seepage relocates dissolved and fine solid substances from the surface to greater depth and from convex terrain to the adjacent gullies and other concavities. Therefore, convexities are usually relatively poor in nutrients [45].
Bedrock outcrops, roche moutonnées and rock bars (Lithic Leptosols; Stöhr 2007) [74] are partly covered with rapidly draining skeletal soil and/or fragmented lichen mats associated with moss layers and dwarf shrubs. Drought, low winter temperatures and limited nutrient availability are hostile to tree establishment in such places. Bedrock clefts, however, occasionally provide suitable conditions for seedlings on such terrain.
Concavities
Due to surface runoff, lateral seepage and snow accumulation, soil moisture and amounts of nutrients increase at the foot of convex slopes and in adjacent gullies and other concavities [45]. Due to the insulation from deep (2-3 m) winter snowpack, soil temperatures do not drop far below the freezing point and rise to zero (0℃) even before the snow melts. Freeze-thaw events are considerably reduced. In snow-rich gullies, decomposition during wintertime may account for 40% of the total annual decomposition [133]. In steep gullies, avalanches and late-lying snow, rather than soil conditions, prevent the long-term establishment of trees. In contrast to steep gullies, that are normally 'well-drained' or 'excessively-drained' [45,105], stagnant water conditions may occur in closed concavities (e.g., kettle holes), on small alluvial flats on gently sloping or level terrain. In both the alpine and the northern treeline ecotone, Gleysols, Stagnosols, and Histosols (peat-land) are found in such very poorly-drained places.
Gradient conditions may be reflected in the distribution pattern of plant communities [8,32,45,123,134]. On gentle trough shoulders (Figure 10) or gently sloping northern fells and in glacial cirques (Figure 11), for example, roche moutonnées associated with former glacial drainage channels, spillways, and small alluvial flats cause a distinctive distribution of vegetation and soil types. At the Rocky Mountain treeline, for example, Dystric Cambisols and also Podzols are found on bedrock outcrops covered with conifers, whereas organic soils developed in the depressions because of poor drainage or even water saturation, deep late-lying winter snowpack, and resultant poor decomposition. Willow thickets, bog and spring communities typically occur in these wet places [31,135,136]. However, the situation in the Alps differs as the anthropogenic upper limit of the treeline ecotone is usually located at lower elevations than the cirque-floors and back-slopes.
The effects of microtopography on microclimates and consequently on soils vary depending on regional climate. They are most remarkable in continental climates with relatively low precipitation, long exposure to sunlight and intense direct solar radiation. Similar effects may occur in climates with a dry summer season. The influence of slope orientation on microclimates and soil pattern is of major importance in this respect (Figure 12 and Figure 13).
Soils in the Treeline Ecotone and Climate Change
Potential effects of changing thermal and humid regimes on soils in the treeline ecotone can be roughly assessed. Recurrent periods of drought, particularly in continental climates, are likely to reduce decomposition and impede root development, and thus nutrient supply. In addition, increasing wild fires would not only destroy trees, but also the organic layer and thereby the main nutrient source.
As soil physical and chemical properties are usually more closely related to the effects of the varying microtopography and geological substrates than to elevational gradients, potential changes need to be assessed at local and regional scales. Thus, in the mountain treeline ecotone on heavily dissected and rugged mountains, stagnant water conditions are restricted to relatively small depressions, and the peat is relatively shallow compared to the vast paludified areas on gently rolling land surfaces in the north. Under warmer and drier conditions, mineralization of these peaty soils (Histosols) will accelerate. Better nutrient availability may facilitate colonization of such sites by trees and shrubs, whereas the convexities soils will be affected by more frequent droughts [3,137,138]. Under the influence of warmer and more humid conditions, on the other hand, decomposition and leaching of soils are likely to increase on convex terrain. Better nutrient availability, however, might be overcompensated by increased leaching.
By contrast, in waterlogged depressions an increase of anaerobic decomposition and release of methane to the atmosphere are likely. Deposition of drifting snow and a greater amount of melt water may play an important role. In addition, wet conditions are adverse for mycorrhizae and thus impede nutrient uptake. In the treeline ecotone on gently rolling land surfaces in the north, such changes would influence much larger areas than in steep-sided mountains.
As may be concluded from snow fence experiments in Alaska tree islands and tree clusters becoming established in the present tundra may locally destabilize permafrost and thermokarst [127]. Thus, a long-term cycle controlled by the interaction of Pinus pumila patches and the permafrost level as described by Kryuchkov [31,139] from the northern treeline in Siberia is not unlikely.
While historical removal of high-elevation forests has had lasting effects on the soils in the treeline ecotone, treeline upward shift - irrespective of whether driven by climate warming and/or cease of pastoral use - brings about changes in soil conditions in areas currently devoid of tree growth. Infilling these gaps and forest in growth [140-146] will probably have a greater effect than establishment of new trees above the existing tree limit. In the Ural Mountains, for example, climatically-driven forest expansion into the mountain tundra is likely to increase ectomycorrhizal biomass [147].
The influence of altered vegetation on litter input and microclimate on soil organic matter may result in a faster cycling of organic matter and increase available nitrogen in the area being invaded by trees [26,68]. In the long-term, however, nitrogen turnover would decrease due to fixation in long-lived trees [148]. Moreover, increasing tree density may buffer warming of the soil and reduce decomposition of the litter. Thus, organic layers would increase and podzolisation may become revived, provided that percolation is sufficient. However, direct (primary) and indirect (secondary) effects of climate warming overlap. Their effects are often difficult to distinguish [26].
Conclusions
No treeline-specific soil types exist. The direct and indirect effects of topographical factors on soil forming processes and vegetation development are key factors in the treeline ecotone and the adjacent alpine zone. Thus, the spatial distribution pattern of soils and the multiple interactions of soil formation processes and vegetation development in the mountain treeline environment can only be assessed at local and regional scales.
In the European Alps, the anthropogenically induced after-effects have a lasting influence on soils in the current treeline ecotone for many hundreds or thousands of years at different intensities. In northern Europe and in the Rocky Mountains, humans have influenced soils more locally. The after-effects of anthropogenic impact may overrule the influence of climate on soil development.
Forest expansion to higher elevations than at present will be associated with changes in soil conditions. Potential variations, however, are difficult to predict, as at small scales (local, micro), climatic (thermal) differences may occur that are as great in magnitude as those that occur over thousands of kilometers in lowland sites. Soil responses to changing climate are a long-term process lagging far behind forest advance. Increasing tree cover, litter supply and reduced decomposition will cause changes in the medium term (decades). Such changes will be more important in the present ecotone than above the existing tree limit. Abrupt changes caused by severe fires, landslides, wind throw and other natural factors may also have lasting effects on soil development in both the medium- and long-term.
References
- Holtmeier FK (1993) Timberlines as indicators of climatic change: Problems and research needs. In: Frenzel B, Eronen M, Vorren KD, et al. Oscillations of the alpine and polar tree limits in the Holocene. Palaeoclimate Research 9, Special Issue: ESF Project "European Plaeoclimate and Man" 4: 211-222.
- Smith WK, Germino MJ, Hancock TE, et al. (2003) Another perspective on altitudinal limits of alpine timberlines. Tree Physiol 23: 1101-1112.
- Holtmeier FK, Broll G (2005) Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Global Ecol Biogeogr 14: 395-410.
- Wieser G, Holtmeier FK, Smith WK (2015) Treelines in a changing global environment. In: Tausz M, Grulke N, Trees in a changing environment: Ecophysiology, Adaptation, and Future Survival. Springer, Berlin, 221-265.
- Neuwinger I (1970) Boden der subalpinen und alpinen Stufe in den Tiroler Alpen. Mitt Ostalpin-Dinarischen Ges Vegetationskunde, Triest 11: 135-150.
- Neuwinger I (1987) Bodenökologische Untersuchungen im Gebiet Obergurgler Zirbenwald Huhe Mut. In: Patzelt G, MaB-Projekt Obergurgl. Veröffentlichungen des Österreichischen MaB-Programms, AUT, 173-190.
- Blaser P, Reiser M (1975) Eine topographische Bodensequenz in subalpinen Lawinengassen auf Silikatgestein. Mitt Eidgenöss Anst forstl Vers wes 51: 1199-1214.
- Blaser P (1980) Der Boden als Standortsfaktor bei Aufforstungen in der subalpinen Stufe (Stillberg, Davos). Schweizerische Anstalt für das Forstliche Versuchswesen, Berichte 56 (3) CH.
- Neuwinger I, Wieser G, Winklehner W, et al. (1988) Bodenwasseruntersuchungen in einer Hochlagenaufforstung bei Haggen im Sellraintal, Tirol. Oesterr Wasser-Abfallwirtsch 40: 57-61.
- Johnson DD, Cline AJ (1965) Colorado mountain soils. Advances in Agronomy 17: 233-281.
- Bockheim JG (1972) Effects of alpine and subalpine vegetation on soil development. University of Washington, USA.
- Retzer JL (1974) Alpine soils. In: Ives JD, Barry RG, Methuen, Arctic and alpine environment. UK, 771-802.
- Burns SF (1980) Alpine soil distribution and development. University of Colorado, USA.
- Burns SF, Tonkin PJ (1982) Soil-geomorphic models and the spatial distribution and development of alpine soils. In: Thorn CE, Allen and Unwin, Space and time in geomorphology. UK, 25-43.
- Litaor MI (1987) The influence of eolian dust on the genesis of alpine soils in the Front Range, Colorado. Soil Sci Soc Am J 51: 142-147.
- Bowman WD, Theodose TA, Schardt JC, et al. (1993) Constraints on nutrient availability on primary production in two alpine tundra communities. Ecology 74: 2085-2097.
- Weiss L, Shiels AB, Walker LR (2005) Soil impacts of bristlecone pine (Pinuslongaeva) tree islands on alpine tundra, Charleston Peak, Nevada. Western North American Naturalist 65: 536-540.
- Robert McNown W, Sullivan PF (2013) Low photosynthesis of treeline white spruce is associated with limited soil nitrogen availability in the western Brooks Range, Alaska. Functional Ecology 27: 672-683.
- Bockheim JG, Munroe JS (2014) Organic carbon pools and genesis of alpine soils with permafrost: A review. Arctic, Antarctic, and Alpine Research 46: 987-1006.
- Romeo R, Vita A, Manuelli S, et al. (2015) Understanding mountain soils: A contribution from mountain areas to the International Year of Soils 2015. FAO, Italy.
- Müller M, Schickhoff U, Scholten T, et al. (2016) How do soil properties affect alpine treelines? General principles in a global perspective and novel findings from Rolwaling Himal, Nepal. Progress in Physical Geography: Earth and Environment 40: 135-160.
- Egli M, Poulenard J (2017) Soils of mountainous landscapes. The International Encyclopedia of Geography.
- Lal R, Kimble JM, Stewart BA (2000) Global climate change and cold regions ecosystems. Lewis Publishers, Boca Raton, Florida.
- Shaver GR, Canadell J, Chapin III FS, et al. (2000) Global warming and terrestrial ecosystems: A conceptual framework for analysis. BioScience 50: 871-882.
- Biasi C, Meyer H, Rusalimova O, et al. (2008) Initial effects of experimental warming in carbon exchange rates, plant growth and microbial dynamics of a lichen-rich dwarf shrub tundra in Siberia. Plant Soil 307: 191-205.
- Kammer A, Hagedorn F, Shevchenko I, et al. (2009) Treeline shifts in the Ural mountains affect soil organic matter dynamics. Global Change Biology 15: 1570-1583.
- Jones A, Stolbovoy V, Broll G, et al. (2010) Soil atlas of the northern circumpolar region. European Commission, Publications Office of the European Union, Luxembourg.
- Chersich S, Rejšek K, Vranova V, et al. (2015) Climate change impacts on the Alpine ecosystem: An overview with focus on the soil -a review. Journal of Forest Science 61: 496-514.
- Mayor JR, Sanders NJ, Classen AT, et al. (2017) Elevation alters ecosystem properties across temperate treelines globally. Nature 542: 91-95.
- Moscatelli MC, Bonifacio E, Chiti T, et al. (2017) Soil properties as indicators of treeline dynamics in view of anthropogenic pressure and climate change. Climate Research 73: 73-84.
- Holtmeier FK (2009) Mountain timberlines. Ecology, patchiness, and dynamics. Advances in global change research.
- Holtmeier FK, Broll G (2010) Wind as an ecological agent at treelines in North America, the Alps, and in the European Subarctic. Physical Geography 31: 203-233.
- Holtmeier FK, Broll G (1992) The influence of tree islands on microtopography and pedoecological conditions in the forest-alpine tundra Ecotone on Niwot Ridge, Colorado Front Range, USA. Arctic and Alpine Research 24: 216-228.
- Broll G, Holtmeier FK (1994) Die Entwicklung von Kleinreliefstrukturen im Waldgrenzökoton der Front Range (Colorado, USA) unter dem Einfluß leewärts wandernder Ablegergruppen (Piceaengelmannii und Abieslasiocarpa). Erdkunde 48: 48-59.
- Schmid GL, Butler DR, Malanson GP, et al. (2009) Soil and pedogenesis at alpine treeline. In: Butler DR, Malanson GP, Walsh SJ, et al., The changing alpine treeline: The example of Glacier National Park, MT, USA, 107-118.
- Thébault A, Clément JC, Ibanez S, et al. (2014) Nitrogen limitation and microbial diversity at the treeline. Oikos 123: 729-740.
- Troll C (1944) Strukturböden, Solifluktion und Frostklimate der Erde. Geologische Rundschau 34: 545-694.
- Washburn AL (1979) Geocryology. A survey of periglacial processes and environments. Edward Arnolds, London, 406.
- Zhang T (2015) Influence of the seasonal snow cover on the ground thermal regime: An overview. Rev Geophys 43: 1-23.
- French HM (2013) The periglacial environment. In: Chichester. (4th edn), John Wiley and Sons Ltd, 544.
- Pisabarro A, Pellitero R, Serrano E, et al. (2016) Ground temperatures, landforms and processes in an Atlantic mountain, Cantabrian Mountains (Northern Spain). Catena 149: 623-636.
- Becker A, Körner C, Brun JJ, et al. (2007) Ecological and land use studies along elevational gradients. Mountain Research and Development 27: 58-65.
- Hiller B, Müterthies A, Broll G, et al. (2002) Investigations on spatial heterogeneity of humus forms and natural regeneration of larch (Larix decidua Mill.) and Swiss stone pine (Pinus cembra L) in an alpine timberline ecotone (Upper Engadine, Central Alps, Switzerland). Geogr Helv Jg 57: 81-90.
- Hiller B, Nuebel A, Broll G, et al. (2005) Snowbeds on silicate rocks in the Upper Engadine (Central Alps, Switzerland)-pedogenesis and interactions among soil, vegetation, and snow cover. Arct Antarc Alp Res 37: 467-476.
- Broll G, Holtmeier FK, Anschlag K, et al. (2007) Landscape mosaic in the treeline ecotone on Mt Rodjanoaivi, Subarctic Finland. Fennia 185: 89-105.
- Broll G (1994) Influence of the soil mosaic on biodiversity at heath sites in the European Subarctic. 15th World Congress of Soil Science, Mexico. 4a: 220-231.
- Loomis OF, Ruess W, Sveinbjörnsson B, et al. (2006) Nitrogen cycling at treeline: Latitudinal and elevational patterns across a boreal landscape. Ecoscience 13: 544-556.
- Haselwandter K (2007) Mycorrhiza in the alpine treeline ecotone: Nutritional implications. In: Wieser G, Tausz M, Trees at their upper limit. Treelife limitation at the alpine timberline, Springer, NL, 57-66.
- Moser M (1982) Mycoflora of the transitional zone from subalpine forests to alpine tundra. Arct Alp Mycol 1: 371-384.
- Kernaghan G, Harper KA (2001) Community structure of ectomycorrhizal fungi across an alpine/subalpine ecotone. Ecography 24: 181-188.
- Geml J (2017) Altitudinal gradients in mycorrhizal symbioses. The current state of knowledge on how richness and community structure change with elevation. In: Tedersoo J, Biogeography of mycorrhizal symbioses. Ecol Stud 230: 107123.
- Holtmeier FK (1974) Geoökologische Beobachtungen und Studien an der subarktischen und alpinen Waldgrenze in vergleichender Sicht (nördliches Fennoskandien, Zentralalpen). Erdwissenschaftliche Forschung VIII. Franz Steiner Verlag, GER.
- Price LW (1969) The collapse of solifluction lobes as a factor in vegetating blockfields. Arctic 22: 395-402.
- Troll C (1973) Rasenabschälung (Turf exfoliation) als periglaziales Phänomen der subpolaren Zonen und der Hochgebirge. Z Geomorphol 17: 1-32.
- Butler DR, Malanson GP, Resler LM, et al. (2009) Geomorphic patterns and processes at alpine timberline. In: Butler DR, Malanson GP, Walsh SJ, et al., The changing alpine treeline: The example of Glacier National Park. Development in Earth Surface Processes, USA, 65-84.
- Burga CA (1988) Swiss vegetation history during the last 18000 years. New Phytologist 110: 581-662.
- Carnelli AL, Theurillat JP, Thinon M, Vadi G, Talon B (2004) Past uppermost tree limit in the central European alps (switzerland) based on soil and soil charcoal. The Holocene 14: 393-405.
- Burga CA, Perret R (2001) Monitoring of eastern and southern Swiss alpine timberline ecotones. In: Burga C, Kratochwil A, Biomonitoring: General and applied aspects on regional and global scale. Springer, Netherlands, 179-194.
- Scheurer T, Jenny H, Kupper P, et al. (2014) Die Landschaft und ihre Nutzung von der letzten Eiszeit bis zur Parkgründung. In: Baur B, Scheurer T, Wissen schaffen, 100 Jahre Forschung im Schweizerischen Nationalpark. Haupt Verlag, Bern, Switzerland.
- Vincze L, Orbán I, Birks H, et al. (2017) The Holocene treeline and timberline changes in the South Carpathians (Romania): Climatic and anthropogenic driver on the southern slopes of the Retezat Mountains. The Holocene 27: 1613-1630.
- Schwörer C, Kaltenrieder P, Glur A, et al. (2014) Holocene climate, fire and vegetation dynamics at the treeline in the northwestern swiss alps. Vegetation History and Archaeobotany 23: 479-496.
- Pini R, Ravazzi C, Raiteri L, et al. (2017) From pristine forests to high-altitude pastures: An ecological approach to prehistoric human impact on vegetation and landscapes in the western Italian Alps. J Ecol 105: 1580-1597.
- Schickhoff U (2005) The upper timberline in the Himalayas, Hindu Kush and Karakorum: A review of geographical and ecological aspects. In: Broll G, Keplin B, Mountain ecosystems. Springer, USA, 274-354.
- Benedict JB (1992) Footprints in the snow: High altitude cultural ecology of the colorado front range, U.S.A. Arctic and Alpine Research 24: 1-16.
- Evans R (1995) Impacts of reindeer grazing in soils and vegetation in Finnmark. NORUT informasjonsteknologi, Rapport 2003/02-94, Northern Research Institute, Tromsö.
- Väre H, Ohtonen R, Mikkola K (1996) The effect and extent of heavy grazing by reindeer in oligotrophic heaths in northern Fennoscandia. In: Väre H, Relationship between spatial heterogeneity in vegetation communities and in belowground ecosystems. Acta Univ Oulu Ser A 4275: 1-14.
- Holtmeier FK, Broll G, Anschlag K (2004) Wind erosion und ihre Folgen im Waldgrenzbereich und in der alpinen Stufe einiger nordfinnischer Fjelle. GEO-öKO 25: 203-224.
- Parker TC, Subke JA, Wookey P (2015) Rapid carbon turnover beneath shrub and tree vegetation is associated with low carbon stocks at a subarctic treeline. Glob Chang Biol 21: 2070-2081.
- Simard M, Lecomte N, Bergeron Y, et al. (2007) Forest productivity decline caused by successional paludification of boreal soils. Ecol Appl 17: 1619-1637.
- Crawford RM, Jeffree CE, Rees WG (2003) Paludification and forest retreat in northern oceanic environment. Ann Bot 91: 213-226.
- Butler DR (1995) Zoogeomorphology: Animals as geomorphic agents. Cambridge University Press, USA.
- Wilkinson MT, Richards PJ, Humphreys GS (2009) Breaking ground: Pedological, geological, and ecological implications of soil bioturbation. Earth-Science Reviews 97: 257-272.
- Holtmeier FK (2015) Animals' influence on the landscape and its ecological importance. Natives, Newcomers, Homecomers. Springer, Netherlands.
- Stöhr D (2007) Soils - heterogeneous at microscale. In: Wieser G, Tausz M, Trees at their upper limit, treelife limitation at the alpine timberline. Plant Ecophysiology 4: 37-56.
- Karlsson PS, Nordell O (1996) Effects of soil temperature on the nitrogen economy and growth of mountain birch seedlings near its presumed low temperature distribution limit. Ecoscience 3: 183-189.
- Smith WK, Germino MJ, Johnson DM, et al. (2009) The altitude of alpine treeline: A bellwether of climate change effects. Bot Rev 75: 163-190.
- Massman W, Sommerfeld R, Zeller K, et al. (1995) CO2 flux through a wyoming seasonal snowpack: Diffusion and pressure pumping effects. In: Tonnessen KA, Williams MW, Tranter M, Biogeochemistry of seasonally snow-covered catchments. IAHS Publ 228: 71-79.
- Brooks PD, Williams MW, Schmidt SK (1996) Microbial activity under alpine snowpacks, Niwot Ridge, Colorado. Biogeochemistry 32: 93-113.
- Jones HG (1999) The ecology of snow-covered systems: A brief overview of nutrient cycling and life in the cold. Hydrol Process 13: 2135-2147.
- Brooks PD, Grogan P, Templer PH, et al. (2011) Carbon and nitrogen cycling in snow-covered environment. Geography Compass 5: 682-699.
- Liptzin D, Hemig D, Schmidt SK, et al. (2015) Winter gas exchange between the atmosphere and snow-covered soils on Niwot Ridge, Colorado, USA. Plant Ecology and Diversity 8: 677-688.
- Eckstein RL (2000) Nitrogen retention by Hylocomium splendens in a subarctic birch woodland. Journal of Ecology 88: 506-515.
- Orlova MA, Lukina NV, Tutubalina OV, et al. (2013) Soild nutrients spatial variability in forest-tundra ecotones on the Kola Peninsula, Russia. Biogeochemistry 113: 283-305.
- Gracanin Z (1972) Die Böden der Alpen. In: Ganssen R, Bodengeographie. Stuttgart, 72-160.
- Compostela C (2011) Paleos uoliedaltriarchivi paleo ambientali per la reconstruzion edell efluttuazion iolocenicedella treeline alpine et appenninica. Università degli Studi di Milano, IT.
- Müller M (1987) Bodenbildung auf Silikatunterlage in der alpinen Stufe des Oberengadins (Zentralalpen, Schweiz). Catena 14: 419-437.
- Bouma J, Hoeks J, Vander Plas L, et al. (1969) Genesis and morphology of some alpine Podzol profiles. J Soil Sci 20: 384-398.
- Haapasaari M (1988) The oligotrophic heath vegetation of northern Fennoscandia and its zonation. Acta Bot Fenn 135: 1-219.
- Jauhiainen E (1969) On soil in the boreal coniferous region. Fennia 98: 1-123.
- Aas B, Faarlund T (2001) The Holocene history of the Nordic mountain birch belt. In: Wielgolaski FE, Nordic mountain birch ecosystems. Man and Biosphere Series 27: 5-22.
- Mahaney WC, Earl J, Kalm V, et al. (1995) Geoecology of the Norra Storfjället area, Northern Sweden. Mt Res Dev 15: 165-174.
- Kullman L, öberg L (2013) Melting glaciers and ice patches in Swedish Lapland provide new insights into the Holocene arboreal history. GEO-öKO 33: 121-146.
- Holtmeier FK, Broll G (2006) Radiocarbon-dated peat and wood remains from the Finnish Subarctic: Evidence of treeline and landscape history. The Holocene 16: 743-751.
- Broll G (2000) Influence of overgrazing by reindeer on soil organic matter and soil microclimate of well-drained soils in the Finnish Subarctic. In: Lal R, Kimble JM, Stewart BA, Global climate change and cold regions ecosystems. Adv Soil Sci 163-172.
- Meier KD, Thannheiser D, Wehberg J, et al. (2005) Soils and nutrients in northern mountain birch forests: A case study from Finnmarksvidda, Northern Norway. In: Wielgolaski FE, Plant ecology, herbivory, and human impact in Nordic mauntain birch forests. Ecol Stud 180: 19-32.
- Thorn CE, Darmody RG (1980) Contemporary eolian sediments in the alpine zone, colorado front range. Physical Geography 1: 162-171.
- De Angelis M, Gaudichet A (1990) Saharan dust deposition over Mont Blanc (French Alps) during the last 30 years. Tellus Series B 43: 61-75.
- Franzén LG, Mattson JO, Martensson U, et al. (1994) Yellow snow over the Alps and subarctic from dust storm in Africa 1991. Ambio 23: 233-235.
- Schwikowski M, Seibert P, Baltensperger U, et al. (1995) A study of an outstanding Saharan dust event at the high-alpine site Jungfraujoch, Switzerland. Atmos Environ 29: 1829-1842.
- Küfmann C (2008) Are cambisols in alpine karst autochtonuous or eolian in origin? Arct Antarc Alp Res 40: 506-518.
- Liu J, Diamond J (2005) China's environment in a globalizing world. Nature 435: 1179-1186.
- Painter TH, Barret AP, Landry CC, et al. (2007) Impact of disturbed desert soils on duration of mountain snow cover. Geophysical Research Letters 34.
- Steltzer H, Landry C, Painter TH, et al. (2009) Biological consequences of earlier snowmelt from desert dust deposition in alpine landscapes. Proc Natl Acad Sci USA 106: 11629-11634.
- Mosimann T (1981) Bodentypen und ihre Verbreitung im Bereich der Waldgrenze am NW-rand der Finnmarksvidda, Norwegen. Nor Geogr Tidsskr 35: 2009-2226.
- Bednorz F, Reichstein M, Broll G, et al. (2000) Humus forms in the forest-alpine tundra ecotone at Stillberg (Dischmatal, Switzerland): spatial heterogeneity and classification. Arctic Antarctic and Alpine Research 32: 21-29.
- Butler DR, Malanson GP, Walsh SJ, et al. (2007) Influences of geomorphology and geology on alpine treeline in the American West - more important than climatic influences? Physical Geography 28: 434-450.
- Malanson GP, Brown DG, Butler DR, et al. (2009) Ecotone dynamics: Invasibility of alpine tundra by tree species from the subalpine forest. In: Butler DR, Malanson GP, Walsh SJ, et al. The changing alpine treeline: The example of Glacier National Park, MT, USA. Development in Earth Surface Processes 12. Elsevier B V, NL, USA, GER, 12: 35-61.
- Holtmeier FK, Broll G (2012) Landform influences on treeline patchiness and dynamics in a changing climate. Physical Geography 33: 403-437.
- Valtera M, Šamonil P, Svoboda M, et al. (2015) Effects of topography and forest stand dynamics on soil morphology in three natural mountain forests. Plant and Soil 392: 57-69.
- Wieder WR, Knowles JF, Blanken PD, et al. (2017) Ecosystem function in complex mountain terrain: Combining models and long-term observations to advance process-based understanding. Journal of Geophysical Research: Biogeosciences 122: 825-845.
- Erickson TA, Williams MW, Winstral A (2005) Persistence of topographic controls on the spatial distribution of snow in rugged mountain terrain, Colorado, United States. Water Resources Research 41.
- Holtmeier FK (2005) Relocation of snow and its effects in the treeline ecotone - with special regard to the Rocky Mountains, the Alps and Northern Europe. Erde 136: 343-373.
- Hellwig N, Anschlag K, Broll G, et al. (2017) Upscaling the spatial distribution of enchytraeids and humus forms in a high mountain environment on the basis of GIS and fuzzy logic. European Journal of Soil Biology 79: 1-13.
- Holtmeier FK, Broll G, Anschlag K, et al. (2003) Regeneration of trees in the treeline ecotone: Northern Finnish Lapland. Fennia 181: 103-128.
- Anschlag K, Broll G, Holtmeier FK (2008) Mountain birch seedlings in the treeline ecotone, Subarctic Finland. Variation in above- and below-ground growth in relation to microtopography. Arctic, Antarctic, and Alpine Research 40: 609-616.
- Turner H (1958) Maximal temperaturen oberflächennaher Bodenschichten an der alpinen Waldgrenze. Wetter Leben 10: 1-12.
- Rien M, Spengler T, Richter M (1998) Klimaökologische Aspekte in Gebirgen der südwestlichen USA unter besonderer Berücksichtigung der White Mountains. Mitteilungen der Fränkischen Geographischen Gesellschaft 45: 303-336.
- Larcher W (1980) Klimastreß im Gebirge, Adaptionstraining und Selektionsfilter für Pflanzen. Rheinische Akademie der Wissenschaften, Vorträge 291: 49-88.
- Ryyppö A, Repo T, Vappaavuori E (1998) Development of freezing tolerance in roots and shoots of scots pine seedlings at non-freezing temperatures. Canadian Journal of Forest Research 28: 557-565.
- Repo T, Nilsson JE, Rikala R, et al. (2001) Cold hardiness of Scots pine (Pinussylvestris). In: Bigras FJ, Colombo SJ, Conifer cold hardiness. Tree Physiol, 1: 463-493.
- Sutinen ML, Ritari A, Holappa T, et al. (2007) Seasonal changes in soil temperature and in the frost hardiness of Scots pine under subarctic conditions. In: Islander IK, Wright AA, Radke JK, et al., International symposium on physics, chemistry and ecology of seasonally frozen soils. Cold Regions Research and Engineering Laboratory, CAN, 513-517.
- Reinhart K, Germino MJ, Kueppers LM, et al. (2015) Linking carbon and water relations to drought-induced mortality in Pinusflexilis seedlings. Tree Physiol 35: 771-782.
- Aulitzky H (1963) Bioklima und Hochlagenaufforstung in der subalpinen Stufe der Inneralpen. Schweizerische Zeitschrift für Forstwesen 114: 1-25.
- Brooks PD, Williams MW, Walker DA, et al. (1995) The Niwot Ridge snow fence experiment: Biogeochemical response to changes in seasonal snowpack. In: Tonnessen KA, Williams MW, Tranter M, Biogeochemistry of seasonally snow-covered catchments. Proc Symp Boulder 1995. International Association of Hydrological Sciences (IAHS), Publication 228, Wallingford, 293-302.
- Edwards AC, Scalenghe R, Freppaz M (2007) Changes in seasonal snow cover of alpine regions and its effects on soil processes: A review. Quaternary International 162-163: 172-181.
- Elliott GP, Cowell LM (2005) Slope aspect mediates fine-scale tree establishment patterns at upper treeline during wet and dry periods of the 20th century. Arctic, Antarctic, and Alpine Research 47: 681-692.
- Hinkel KM, Herd JK (2006) Permafrost destabilization and thermokarst following snow fence installation, Barrow, Alaska, USA. Arctic, Antarctic, and Alpine Research 38: 530-539.
- Li X, Luo T (2011) Spatiotemporal variability of soil temperature and moisture in the south eastern Tibetan Plateau after 200 years warming. New Phytol 19: 760-769.
- Marr W (1966) The development and movement of tree islands near the upper limit of tree growth in the southern Rocky Mountains. Ecology 58: 1159-1164.
- Turner H, Häsler R, Schönenberger W (1982) Contrasting microenvironments and their effects on carbon uptake and allocation by young conifers near alpine treeline in Switzerland. In: Waring RH, Proceedings IUFRO workshop. Ecology of Subalpine Zones, Corvallis, Oregon, United States, 22-30.
- Alvarez-Uria P, Körner C (2007) Low temperature limits of root growth in deciduous and evergreen temperate tree species. Functional Ecology 21: 211-218.
- Turner H, Streule P (1983) Wurzelwachstum und Sprossentwicklung junger Koniferen im Klimastress der alpinen Waldgrenze, mit Berücksichtigung von Mikroklima, Photosynthese und Stoffproduktion. In Wurzelökologie und ihre Nutzanwendung. Int. Symp. Gumpenstein 1982: Bundesanstalt Gumpenstein, Irdning, 617-635.
- Reichstein M, Bednorz F, Broll G, et al. (2000) Temperature dependence of carbon mineralisation: Conclusions from a long-term incubation of subalpine soil samples. Soil Biology and Biochemistry 32: 947-958.
- Aulitzky H (1963) Grundlagen und Anwendung des vorläufigen Wind-Schnee Ökogramms. Mitt Forstl Bundesversuchsanstalt Mariabrunn 60: 763-834.
- Komarkova V (1979) Alpine vegetation of the Indian Peaks area, Front Range, Colorado Rocky Mountains. Flora et Vegetatio Mundi 1: 547-591.
- Haase R (1987) An alpine vegetation map of Caribou Lake Valley and Fourth of July Valley, Front Range, Colorado, U.S.A. Arctic and Alpine Research 19: 1-10.
- Kullman L (2009) Recent change of alpine vegetation and plant species richness in the Swedish Scandes. Mountain Forum Bulletin 9: 20-21.
- Kullman L (2010) One century of treeline change and stability - an illustrated view from the Swedish Scandes. Landscape Online 17: 1-42.
- Kryuchkov VV (1973) The effect of permafrost on northern treeline. Proc Nat Acad Sci 136-138.
- Szeicz JM, MacDonald GM (1995) Recent white spruce dynamics at the subarctic alpine treeline of north-western Canada. Journal of Ecology 83: 873-885.
- Klasner FL, Fagre DB (2002) Half a century of change in alpine patterns at Glacier National Park, Montana, U.S.A. Arctic, Antarctic, and Alpine Research 34: 49-56.
- Bolstadt PV, Vose JM (2005) Forest pasture, carbon pools and soil respiration in the southern Appalachian Mountains. Forest Science 51: 372-383.
- Shiyatov SG, Terent'ev MM, Fomin V (2005) Spatiotemporal dynamics of forest-tundra communities in the polar Urals. Russian Journal of Ecology 36: 69-75.
- Gehrig-Fasel J, Guisan A, Zimmermann N (2007) Treeline shifts in the Swiss Alps: Climate change or land abandonment? Journal of Vegetation Science 18: 571-582.
- Bryn A (2008) Recent forest limit changes in southeast Norway: Effects of climate change or regrowth after abandoned utilisation? Norwegian Journal of Geography 62: 251-270.
- Feurdean A, Gałka M, Tanţău I, et al. (2016) Tree and timberline shifts in the northern Romanian Carpathians during the Holocene and the response to environmental changes. Quaternary Science Reviews 134: 100-113.
- Solly EF, Dukic I, Moiseev PA, et al. (2017) Treeline advances and associated shifts in the ground vegetation alter fine roots dynamics and mycelia production in the Southern and Polar Urals. Oecologia 183: 571-586.
- Dawes MA, Schleppi P, Hättenschwiler S, et al. (2017) Soil warming opens the nitrogen cycle at the alpine treeline. Global Change Biology 23: 421-434.
Corresponding Author
Gabriele Broll, Institute of Geography, University of Osnabrück, Seminarstrasse 19, D-49074 Osnabrück, Germany, Tel: 0049-541-996-4073.
Copyright
© 2018 Holtmeier FK, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.